Djurfakta

- Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?
- Förnuft, känslor och beteenden – Hur funkar hjärnan, nervsystemet, sinnen och hormoner?
- Att hålla sig varm när det är kallt, och kall när det är varmt – Om djurens kroppstemperaturreglering
- Djurens evolution och människans ursprung – Åldrande och livslängd
- Märkliga och farliga djur
- Varför är vissa djur stora medan andra är små?
- Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
- Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten
- Njurar och utsöndring – Om salt- och vattenbalansen i kroppen
Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?

Varför har vi två näsborrar? Naturligtvis har det att göra med att kroppen har en vänstersida och en högersida, som till största delen är spegelbilder av varandra. Det är därför som det finns ett par av de flesta organ i kroppen, dock inte av alla. Vi har bland annat två näsborrar och näshålor, två ögon, två öron, två lungor och två njurar. Men det är inte hela historien. Näsborrarnas och näshålornas evolutionära historia är mycket intressant och deras funktioner har förändrats under årmiljonernas gång.
Vi skulle kunna säga att vi har två näsborrar dels för att fiskar för mycket länge sedan anpassades till att bättre kunna orientera sig med hjälp av lukten, dels för att ett par näsborrar vandrat in i munhålan. Det här kräver förklaringar. Med näsborrar menar jag här bara mynningar på huvudets utsida, så kallade yttre näsöppningar.
Fiskarnas luktgropar: en eller fyra näsborrar
De flesta fiskar andas genom munnen. De har ingen riktig nos eller näsa att andas genom. Men framför ögonen har de flesta fiskar ett par fördjupningar, en på vardera sidan av huvudet. Dessa fördjupningar kallas luktgropar och är fiskarnas luktorgan. Hos de flesta fiskar sitter de på huvudets översida. Hos hajarna sitter de på undersidan.
De käklösa fiskarna, pirålarna och nejonögonen, har dock ett enda oparigt luktorgan i form av en grop mitt på huvudets ovansida. Pirålarna har dessutom en inre näsöppning, som förbinder näsgropen med svalget, så att vatten kan strömma förbi luktslemhinnan. De käklösa fiskarna är de nutida djur som står närmast ryggradsdjurens, och därmed människans, urfader. Därför är det ingen orimlig tanke att vi skulle kunna härstamma från ett urryggradsdjur med en enda näsborre.
Det är emellertid bra för fiskar att ha två luktorgan, för då kan de lättare avgöra varifrån en lukt kommer. Om det luktar föda i det vänstra luktorganet, men inte i den högra, så simmar fisken åt vänster och hittar födan. De flesta fiskar har fyra näsborrar, en främre och en bakre på varje sida av huvudet. Mellan den främre och den bakre näsborren löper en kanal. När fisken simmar strömmar vattnet in genom den främre näsborren, passerar genom kanalen, där lukten registreras av luktslemhinnan, och strömmar sedan ut genom den bakre näsborren.
Fyrfotadjur: två näsborrar, men fyra näsöppningar
För flera hundra miljoner år sedan uppkom hos en del fiskar en förbindelse mellan luktgroparna och munhålan, de första inre näsöppningarna (också kallade koaner). Alla landlevande ryggradsdjur härstammar från sådana fiskar. Inre näsöppningar finns därför kvar hos alla landryggradsdjur, det vill säga hos groddjur, kräldjur inklusive fåglar och däggdjur. Alla dessa djur har två näsborrar, som också kallas yttre näsöppningar. Har du inre näsöppningar, så behöver du inte andas genom munnen, utan kan i stället andas genom två näsgångar. Dessa gångar börjar med näsborrarna, det vill säga de yttre näsöppningarna, och slutar med de inre näsöppningarna i munhålans tak eller i svalget. Detta är en mycket viktig anpassning till landliv. Det gör nämligen att luftandande djur kan spara på vatten. Andas du genom munnen, avdunstar mycket mer vatten från luftvägarna, än då du andas genom näsgångarna.
Hos groddjur och de flesta reptiler, däribland fåglarna, är näsgångarna utvidgade till näshålor. I de båda hålornas väggar finns luktsinnescellerna. Människor och andra däggdjur har utvecklat en ny skiljevägg, gommen (egentligen den sekundära gommen), som skiljer näshålorna från munhålan. Därigenom har de inre näsöppningarna förskjutits bakåt. Både näshålorna och munhålan mynnar därför baktill i svalget. Gommen är munhålans tak och näshålornas golv. Även krokodiler har utvecklat en sekundär gom.
Inre näsöppningar uppträdde först hos så kallade tetrapodomorfa fiskar för mer än 380 miljoner år sedan. Det var 30 miljoner år innan sådana fiskar började utveckla gångben, gå upp på land och utvecklas till de första groddjursliknande landryggradsdjuren. Detta anses tyda på att de inre näsöppningarnas ursprungliga funktion inte hade med luftandning att göra. Under dessa fiskars evolution vandrade de bakre näsborrarna ner mot överkäkens underkant, passerade genom tandraden och hamnade i muntaket som inre näsöppningar. De var de första ryggradsdjuren med två näsborrar. Forskare har hittat ett fossil som är en perfekt övergångsform ("felande länk") med de bakre näsborrarna i överkäkens kant.
"Falska" näsöppningar hos fiskar
Förbindelser mellan luktgroparna och munhålan har också utvecklats hos några äkta benfiskar (Teleostei) som en anpassning till ett nedgrävt levnadssätt. Dessa fiskar är inte närmare släkt med landryggradsdjurens förfäder. Till dem hör vissa stjärnkikare (släktet Astroscopus). Deras förbindelse är en nybildning, alltså en "falsk" inre näsöppning.

Lungfiskar: fyra näsöppningar men inga näsborrar
Hos den gren av fiskarnas stamträd som lett till lungfiskarna har bakre näsöppningar vandrat in i munhålan och mynnar där, medan de främre är belägna i anslutning till överläppen och mynnar i den främre delen av munhålan. Eftersom båda paren av näsöppningar mynnar i munhålan, skulle vi kunna säga att lungfiskarna inte har några näsborrar alls. De flesta menar att lungfiskarna utvecklat detta arrangemang parallellt med och oberoende av de fiskar som gav upphov till landryggradsdjuren. Detta tyder på att att de fyrfota landryggradsdjurens urfader inte var en lungfisk. Men det finns andra data som tyder tyder på att lungfiskarna är landryggradsdjurens närmaste nu levande släktingar, närmare släkt med dem än tofsstjärtsfiskarna (kvastfeningarna).
Läpp-, käk- och gomspalt: missbildningar som återspeglar vårt evolutionära förflutna
Hos människa är läpp-, käk- och gomspalt relativt vanliga medfödda missbildningar. De beror på att de tre ansiktsdelar som bildar överkäken inte växer ihop under fosterstadiet. När en sådan missbildning är dubbelsidig och fullt utvecklad så återspeglar de båda spalterna precis den väg som fiskarnas bakre näsborrar vandrade tills de blev inre näsöppningar. En evolutionär process som skedde för nästan 400 miljoner år sedan kan alltså ha betydelse för människan än i dag. Numera kan dessa missbildningar korrigeras kirurgiskt med goda resultat.
Referenser
- J. W. Atz: Narial breathing in fishes and the evolution of internal nares (The Quarterly Review of Biology 27:366-377, 1952.
- G. Bertmar: The olfactory organ and upper lips in Dipnoi. an embryological study (Acta Zoologica 46:1-40, 1965)
- P. Janvier: Wandering nostrils (Nature 432:23-24, 2004).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- A.S. Romer: The vertebrate body (3rd ed, Saunders, 1962).
- M. Zhu och P. E. Ahlberg: The origin of the internal nostril of tetrapods (Nature 432:94-97, 2004).

Många djur kan gå och springa med hjälp av sina ben. Men varför har olika djur olika antal gångben? Varför har människor två ben, hästar fyra, myror sex och spindlar åtta gångben? I en del fall kan det bero på anpassningar till miljön, i andra fall skulle det kunna förklaras av djurens evolutionära historia och i vissa fall kan vi inte förklara det.
De flesta djur är bilateralsymmetriska. Detta innebär att de har
- en högersida och en vänstersida som är i det närmaste spegelbilder av varandra
- en framände med huvud och en bakände
- en ryggsida och en buksida
Fördelen med detta är att det blir lätt att röra sig mot ett mål, till exempel mot ett ställe där det finns mat. Det innebär också att djur har lika många gångben på vänster och höger sida av kroppen.
Om däggdjurs och fåglars gångben och om hur de kan få nya funktioner
Vi människor är förstås bilateralsymmetriska. Det är därför vi har två armar, en på varje sida, och två gångben, ett på varje sida. Precis som vi, så har alla djur som går med ben ett jämnt antal ben, lika många på varje sida. Annars hade det blivit svårare att hålla balansen. Fåglar har två gångben. Nästan alla däggdjur har fyra gångben. Bland undantagen hittar vi bland annat människor, kängurur och en del gnagare, som alla bara använder bakbenen när de förflyttar sig.
Hos många av de djur som förflyttar sig på bakbenen har frambenen fått en annan nyttig funktion. Fåglarnas framben har utvecklats till vingar. Eftersom människans framben inte är gångben, kan hon bära tunga saker och använda händerna även när hon rör sig.

Kängururnas framben är däremot mer eller mindre förkrympta och till liten nytta. Varför använder de bara ett par gångben när de rör sig snabbt? Den sannolika förklaringen är att de har utvecklat ett sätt att förflytta sig, som är mindre energikrävande än att springa på fyra ben. Kanske gällde detta också för de tvåbenta, icke flygande dinosaurierna. Tyrannosaurus rex hade enormt förkrympta framben, som inte kan ha varit särskilt användbara till någonting. Även människans bakben har utvecklats till effektiva redskap för långdistanslöpning. Det är därför som vi har de konstigaste fötterna bland alla apor.
Ryggradslösa djurs gångben
Insekter har sex gångben, medan spindlar, dolksvansar och andra spindeldjur samt många kräftdjur har åtta gångben. Mångfotingar ("tusenfotingar") har ett mycket stort antal gångben, dock ej tusen.
Om djur har väldigt många gångben, kan det troligen bli svårt att koordinera rörelserna. Detta kan leda till att de alltid har samma rörelsemönster och har svårt att anpassa rörelserna till nya situationer. Insekter, spindeldjur och kräftdjur med få gångben härstammar från djur med betydligt fler. Genom att minska antalet gångben har dessa djur skaffat sig en betydligt större rörelserepertoar. Detta är troligen en anledning till att dessa djurgrupper är mycket större och artrikare än mångfotingarna.

Varför har landlevande ryggradsdjur fyra ben?
Varför har nästan alla landlevande ryggradsdjur fyra ben, inte två, inte sex och inte åtta? Hos en del av dessa djur fungerar frambenen inte längre som gångben. Hos andra har ett eller båda benparen försvunnit. Valarna har förlorat bakbenen, medan frambenen har omvandlats till fenor. Ormar har förlorat alla benen. Det finns groddjur och ödlor utan ben eller med bara ett par ben. Men alla landryggradsdjur härstammar från djur som gick på fyra ben och inga av dem har fler än fyra ben. Orsaken hittar vi i deras evolutionära historia.
Alla landryggradsdjur härstammar från utdöda fiskar som hade två par fenor längs med sidorna på buken, ett par bröstfenor och ett par bukfenor. Dessa fenor utvecklades till de fyra ben som vi ser hos groddjur, kräldjur inklusive fåglar och däggdjur. Den evolutionära förklaringen till att landryggradsdjuren fick fyra ben skulle således kunna vara att fiskars simning underlättas av att de har just två par fenor på buksidan, inte bara en stjärtfena och opariga ryggfenor. Kanske finns det under embryots utveckling någon slags restriktion, som gör det omöjligt för fyrfota landryggradsdjur att utveckla fler benpar. Sådana restriktioner (på engelska "developmental bias") kan i många fall begränsa de evolutionära möjligheterna för djur.
Vilket är det bästa antalet gångben att ha?
Som ovan nämnts, är det svårt att samordna ett stort antal gångben. Vi ser därför en tendens i djurvärlden till att djur, som är anpassade till en effektiv gång, har ett litet antal gångben. Vilket är då det bästa antalet ben du kan ha, när du rör sig? En hel del talar för att det är sex ben. Insekterna skulle alltså vara bättre konstruerade än vi däggdjur avseende benantal. Det minsta antalet stödpunkter som kan ge stöd åt kroppen utan att du behöver balansera är tre. Tre ben ger dessutom stöd i alla lägen, så länge kroppens tyngdpunkt befinner sig ovanför den triangel som bildas av fötternas stödpunkter på marken. En trebent pall vickar aldrig. De sexbenta insekterna har två uppsättningar med tre ben och kan alltså vid gång växla mellan två mycket stabila lägen. Insekterna har således det minsta antalet gångben som kan ge maximal stabilitet.

Med två eller fyra ben måste djur hålla balansen
Fyrbenta djur är stabila när de står stilla på fyra ben, men när de rör sig vilar de tidvis på ett, två eller tre ben. Vid vissa gångarter ingår dessutom moment då djuret inte vidrör marken. Vid alla gångarter måste djuren balansera kroppen så att de inte faller. Undantag utgör bland annat många groddjur och ödlor, som tar sig fram med buken i kontakt med marken, ofta slingrande som ormar.

Gångarter hos fyrfota djur
Vid långsam gång (hos hästar skritt) har fyrfotadjur alltid tre ben i marken samtidigt. När de rör sig snabbare går de över i trav. Då flyttar de diagonalt motsatta ben nära nog samtidigt: först vänster fram och höger bak, sedan höger fram och vänster bak och så vidare. Periodvis svävar de då ofta i luften, utan att något ben vidrör marken. Vid passgång förflyttas samma sidas ben samtidigt: först höger fram och höger bak, sedan vänster fram och vänster bak. Passgång är ovanligt, men förekommer hos kameler och giraffer samt hos vissa hästraser. Möjligen är det svårare att hålla balansen vid passgång än vid trav, men lättare att undvika att benen kolliderar med varandra. När de rör sig mycket snabbt, använder många däggdjur galopp. Vid hästars galopp sätts aldrig mer än två ben i marken, tidvis bara ett. Dessutom ingår ett moment då djuret svävar i luften utan markkontakt. Många mindre däggdjur rör sig framåt hoppande med fyra ben, så kallat språng. Då vidrör först bakbenen och sedan frambenen marken, i båda fallen nästan samtidigt. Bakbenen utvecklar den största kraften. Vissa djur kan röra sig genom att hoppa med båda bakbenen. Exempel är många grodor, kängurur och vissa smågnagare samt många fåglar. Slutligen har vi tvåbenta djur, som går eller springer genom att omväxlande röra de båda benen, till exempel fåglar och människor.

Människans anpassningar till tvåbenthet
Människor måste alltid balansera kroppen så att dess tyngdpunkt befinner sig ovanför den yta som begränsas av fötternas utsidor. Detta gäller när vi står, när vi går och när vi springer. Annars ramlar vi i sidled, framåt eller bakåt. Vi har emellertid en uppsättning fantastiska reflexer, som håller tyngdpunkten inom detta område. Vi har också skyddsreflexer som gör att vi inte faller, om tyngdpunkten hamnar utanför detta område. Ofta flyttar vi då automatiskt ena foten i fallets riktning och stödjer oss på den. Men vi drattar ändå rätt ofta omkull.
Skillnader mellan att gå och att springa
Hos människan innebär gång att vi först höjer kroppen uppåt genom att skjuta ifrån med det ena benet. Uppåtrörelsen innebär att kroppen får en högre lägesenergi. Sedan faller vi framåt och utnyttjar då lägesenergin genom att omsätta den till rörelseenergi. Samtidigt pendlar det andra benet framåt, sätts i marken och stoppar fallet. Och så håller vi på, hela tiden utnyttjande lagrad lägesenergi. Det är som att resa upp ett ägg på den ena änden, låta det falla framåt och ständigt upprepa manövern.
När vi springer skjuter vi ifrån med det ena benet, så att vi flyger framåt, utan att vidröra marken. Sedan pendlar det andra benet framåt, sätts i marken och stoppar rörelsen. Vid nedslaget omvandlas den rörelseenergi vi skaffat oss vid avstampet till elastisk lägesenergi i det ben som slår i marken. Lägesenergin lagras i spända senor och andra elastiska delar av rörelseapparaten. Denna elastiska energi ger en del av kraften vid nästa avstamp. Och så håller vi på, hela tiden återvinnande en del av rörelseenergin.
Fördjupning
Läs gärna nästa artikel nedan om varför ryggradsdjur aldrig har fler än fem fingrar och tår, men ofta färre?
Referenser
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- G. M. O. Maloiy, B. M. Rugangazi, och M. F. Rowe: Energy expenditure during level locomotion in large desert ungulates: the one-humped camel and the domestic donkey (Journal of Zoology 277:248-255, 2009).
- P. Meurling: Gångart, de fyrfota djurens rörelsemönster (Nationalencyklopedin, hämtad 2022-07-08).

De första ryggradsdjur, som kunde ta sig upp på land, levde under devonperioden (cirka 416-359 år före nutid). De härstammade från fiskar med skaftade fenor. Dessa fiskar utvecklades till olika typer av groddjursliknande djur, hos vilka de pariga fenorna omvandlats till två par gångben. Bland dem fanns det djur med sex, sju eller åtta tår på fötterna. Men det var djur med fem tår på alla fyra fötterna som utvecklades till de nu levande fyrfota ryggradsdjuren: groddjur, reptiler inklusive fåglar samt däggdjur. Varför det?
Ett möjligt undantag är de nutida groddjuren, som har fyra tår på framfötterna och fem tår på bakfötterna. De kan ha utvecklats från en utdöd groddjursgrupp med samma tåantal, fyra fram och fem bak. Mot detta talar det faktum att vissa groddjur i embryot utvecklar ett anlag till en femte framfotstå. Detta anlag försvinner emellertid sedan utan att ge upphov till en tå.

Varför aldrig mer än fem fingrar och tår?
Varför finns det högst fem fingrar och tår hos nutida fyrfotadjur? Det kan ha varit ren slump, som hävdats av den berömde evolutionsforskaren Stephen Jay Gould. Andra har hävdat att det fanns fördelar med fem tår per fot och att detta tåantal gynnades av evolutionens naturliga urval. Klart är i varje fall att inga nutida fyrfotadjur utvecklat fler än fem tår under evolutionens gång. Forskarna trodde tidigare att några av de utdöda fiskliknande ichthyosaurierna ("fisködlorna") var ett undantag. Nyare studier tyder dock på att de inte hade mer än fem tår inuti sina fenor, men att de talrika tåbenen och andra handben inte låg i tydliga räta rader. Vissa tår var också tvekluvna.
Ett möjligt undantag med sex tår är dock klogrodorna (släktet Xenopus). Dessa grodor har framför bakfotens stortå ett kloförsett utskott, som tolkats som en "falsk" stortå, prehallux. I en studie från 2015 tolkas dock detta utskott som en äkta tå och klogrodorna har i så fall möjligen utvecklat en sjätte tå.

Däremot har tåantalet reducerats hos många grupper av fyrfotadjur. Fåglar har fyra tår på bakbenen. Bland annat nötkreatur, får, getter, antiloper och svin har fyra tår på alla fyra benen, av vilka de yttersta (lättklövarna) är mer eller mindre tillbakabildade. Kameldjur har två tår på alla fyra benen. Hästar har bara en tå på alla fyra benen.
De flesta seriefigurer har bara fyra fingrar. Bakgrunden torde inte vara biologisk. Det är mycket lättare att rita händer med färre fingrar.
Missbildningar med fler än fem fingrar eller tår
Extra fingrar eller tår, så kallad polydaktyli, är en inte ovanlig missbildning hos fyrfotadjur, även hos människor. Mot den bakgrunden är det märkligt att antalet fingrar och tår inte utökats under evolutionen. Polydaktyli innebär dock inte att det bildas en ny finger eller en ny tå med unika egenskaper. I stället finns bildas "tvillingar" av redan befintliga fingrar eller tår, till exempel två ringfingrar på samma hand eller två stortår på samma fot. Extratår och extrafingrar kan därför inte modiferas genom naturligt urval, så att handen eller foten genetiskt anpassas till nya funktioner.

Fosterutvecklingen begränsar evolutionen
Evolutionen har således aldrig lett till att fler än fem fingrar och tår utvecklats, med klogrodorna som möjligt undantag. Orsaken skulle kunna vara att embryonalutvecklingens komplexa schema inte tillåter detta.
Anlagen till armar och ben, extremitetsknopparna, utvecklas under en kritisk period i början av embryots utveckling, utom hos groddjuren hos vilka de uppträder senare i samband med metamorfosen från larv till vuxet djur. Under denna kritiska period byggs kroppens grundplan upp. Denna process styrs av en rad proteiner och andra ämnen. Många av dessa ämnen bildas i en del av embryot, transporteras till en annan del och påverkar utvecklingen där. Andra verkar inom extremitetsknopparna och styr deras utveckling till ben eller armar. Ofta är dessutom ett och samma ämne nödvändigt för normal utveckling i flera delar av embryot. Mutationer som ger upphov till nya fingrar eller tår stör sannolikt det intrikata samspelet mellan andra delar av embryot, något som ger skadliga effekter i andra delar av extremiteterna eller i helt andra kroppsdelar. Sådana effekter leder till minskad livsduglighet och försämrad överlevnadsförmåga. Polydaktyli hos människan åtföljs faktiskt ofta av andra fostermissbildningar och kanske också av en ökad risk att drabbas av vissa sjukdomar.
Flera studier tyder på att det är samspelet mellan de så kallade Hox-generna och mellan dem och andra gener, som begränsar antalet fingrar och tår. En särskilt viktig gen är sonic hedgehog, uppkallad efter en figur i ett datorspel. Alla dessa gener måste aktiveras efter ett bestämt mönster i tid och rum, så att de proteiner de kodar för styr embryots utveckling i normal riktning. Annars störs hela embryots utveckling, inte bara utveckling av fingrar och tår.

Falska fingrar och tår
I flera fall har emellertid andra strukturer, som inte är fingrar eller tår, tillkommit under evolutionens gång. De fungerar som extra fingrar eller tår utan att till sitt ursprung vara det.
Den mest berömda "falska tån" är "pandans tumme", egentligen förtumme eller prepollux, diskuterad av Stephen Jay Gould. Jättepandans tumme är inte ett extra sjätte finger, utan en förtjockad trampdyna. Den fungerar däremot som en motsättlig tumme och kan pressas mot handflatan, när pandan handskas med sin favoritföda, bambuskott. Pandans tumme är förstärkt av ett skelettben. Men detta ben är inte ett fingerben, utan ett omvandlat så kallat sesamben. Sesamben är små ben som finns inuti senor, utan kontakt med det övriga skelettet. De har liknats vid sesamfrön, därav namnet. Människans största och mest välkända sesamben är knäskålen.

Hos många mullvadsarter förekommer ett skelettben utanför tummen, en förtumme (prepollux). Det fungerar som ett sjätte mellanhandsben och gör handflatan bredare, något som underlättar grävandet. Men det härstammar inte från ett äkta finger. Det har, precis som pandans tumme, utvecklats ur ett sesamben.
Elefanter har en kraftig broskstav inuti alla fyra fötterna utanför "stortårnas" mellanfotsben. Staven förstärker foten, något som kan vara nödvändigt hos ett så tungt djur. Den är inte ett sjätte mellanfotsben tillhörande en försvunnen sjätte tå. Men den fyller samma funktion som ett mellanfotsben. Staven är således helt jämförbar med pandans och mullvadarnas förtummar. Den har också, liksom dessa, utvecklats ur ett sesamben.
Referenser
- P. E. Ahlberg, J. A. Clack och H. Blom: The axial skeleton of the Devonian tetrapod Ichthyostega (Nature 437:137-140, 2005).
- M. I. Coates och J. A. Clack: Polydactyly in the earliest known tetrapod limbs (Nature 347:66-69, 1990).
- F. Galis, J. J. M. van Alphen och J. A. J. Metz: Why five fingers? Evolutionary constraints on digit numbers (Trends in Ecology & Evolution 16:637-646. 2001).
- S. Hayashi et al.: Evidence for an amphibian sixth digit (Zoological Letters 1:17, 2015).
- Y. Hu et al.: Comparative genomics reveals convergent evolution between the bamboo-eating giant and red pandas (Proceedings of the National Academy of Sciences 114:1081-1086, 2017).
- J. R. Hutchinson et al.: From flat foot to fat foot: structure, ontogeny, function, and evolution of elephant “sixth toes” (Science 334:1699-1703, 2011).
- Y. Kherdjemil et al.: Evolution of Hoxa11 regulation in vertebrates is linked to the pentadactyl state (Nature, doi:10.1038/nature19813, 2016).
- C. Mitgutsch et al.: Circumventing the polydactyly 'constraint': the mole's 'thumb' (Biology Letters 8:74-77, 2012).
- J. Qiu: How the panda’s ‘thumb’ evolved twice (Nature News, 16 januari 2017).
- Aditya Saxena et al.: The origins, scaling and loss of tetrapod digits (Philosophical Transactions of the Royal Society B 372:20150482, 2017).
- G. E. Weissengruber et al.: The structure of the cushions in the feet of African elephants (Loxodonta africana) (Journal of Anatomy 209:781-792, 2006).

Hos människan finns vissa strukturer som inte har någon funktion alls eller en försumbar funktion. Dessa rudiment utgör evolutionära rester, som vi bär med oss utan att de har några väsentliga negativa effekter. Rudiment kan endast förklaras genom evolution med naturligt urval. Exempel på rudiment hos människor är bland annat svansbenet, hårresarmusklerna, ytteröronens muskler och visdomständerna.
Rudiment och exaptioner
Allmänt kan vi säga att en struktur eller en egenskap för det mesta är adaptiv, det vill säga gynnad av det naturliga urvalet, men den behöver inte vara det. Icke funktionella egenskaper eller strukturer kan bland annat finnas på grund av att de etablerats slumpmässigt, på grund av att de behövs för en normal embryonalutveckling eller på grund av att de inte påverkar vår överlevnad och reproduktion. I vissa fall har egenskapen eller strukturen varit funktionell hos våra evolutionära förfäder, men är det inte längre eller har mycket liten betydelse. Då talar vi om rudiment. Rudiment har ingen eller obetydlig negativ effekt på individens livsduglighet och har därför inte sorterats bort av det naturliga urvalet. Strukturer som har förlorat sin gamla funktion, men fått en ny funktion. brukar kallas exaptioner. I likhet med rudiment, kan exaptioner bara förklaras med hjälp av evolution genom naturligt urval.

Det finns många väl belagda exempel på exaptioner hos djur. Fåglarnas vingar och valarnas bröstfenor hat utvecklats ur gångben. Människans fot kan i viss mån betraktas som en exaption. Hos andra apor är den en gripfot, även om den också kan användas vid gång. Hos oss har den förlorat gripfunktionen och adapterats till gång och löpning, samma funktion som den hade hos apornas avlägsna förfäder. Rudiment är svårare att belägga än exaptioner. Det är nämligen alltid svårare att påvisa brist på funktion än att påvisa funktion.

Blindtarmens maskformiga bihang och gommandlarna
Blindtarmens maskformiga bihang (appendix vermiformis) betraktas ofta som ett rudiment utan funktion, men så är det förmodligen inte. Du klarar dig i och för sig bra utan det, undertecknad förlorade sitt år 1952. Bihanget kan ju bli infekterat av farliga bakterier, något som mycket ofta ledde till döden innan kirurger kunde operera bort det. Detta leder till tanken att det skulle ha någon funktion, så att risken att drabbas av appendicit ("blindtarmsinflammation") uppvägs av positiva effekter på överlevnaden.
Det maskformiga bihanget innehåller rikligt med immunologiskt aktiv vävnad. Kanske har det viktiga immunlologiska funktioner under de tidigaste barnaåren. Enligt en intressant hypotes med visst stöd skulle bihanget fungera som en tillflyktsort för tarmens nyttiga mikroorganismer vid tarminfektioner, då dessa organismer sköljs bort från övriga delar av mag-tarmkanalen av diarréer. Därmed skulle tarmen hos dem som överlevt en infektion snabbare kunna koloniseras av en normal tarmflora, som skyddar mot framtida infektioner.
Gomtonsillerna (halsmandlarna) innehåller, precis som det maskformiga bihanget, immunologiskt aktiv vävnad. Detsamma gäller flera andra tonsiller, som tillsammans med gomtonsillerna bildar en ring kring svalgets öppning. Dessa organ har ansetts ha försumbara funktioner, nästan betraktats som rudiment. En del av dem, särskilt gomtonsillerna, opereras ofta bort hos barn på grund av upprepade infektioner eller störd andning, dock mindre ofta numera än tidigare. Forskare har gjort en omfattande studie av barn som fått tonsiller bortopererade före 9 års ålder, jämfört dem med en kontrollgrupp och följt upp dem senare i livet. Forskarna fann en klart ökad risk för sjukdomar i luftvägarna och en viss ökad risk för allergier och infektionssjukdomar hos dem som opererats. Fler studier behövs, men dessa organ har sannolikt viktiga funktioner. Tonsillerna är större hos barn än hos vuxna. Kanske har de funktioner i samband med immunsystemets mognad under barnaåren.
Bröstvårtor hos män
Bröstvårtor och bröstkörtlar hos män fyller ingen funktion och har betraktas som en slags rudiment. Men detta stämmer inte. Bröstvårtor och bröstkörtlar har sannolikt aldrig haft någon funktion hos däggdjurshanar. Ett undantag är storfladdermusen Dyacopterus spadiceus, hos vilken hanarna uppges ge ungarna mjölk. Anlagen till bröstvårtor och bröstkörtlar bildas i fostret, innan det diffferentierats i manlig eller kvinnlig riktning. Det har sannolikt inte varit någon större evolutionär nackdel att behålla dem hos män. Alternativt skulle fosterutvecklingen kanske inte kunna fungera normalt utan dem. Mot den senare hypotesen talar det faktum att nästan alla pungdjurshanar saknar bröstvårtor. Förekomsten av prostatavävnad hos kvinnor kan förklaras på ett liknande sätt. De två gångsystem som ingår i de kvinnliga respektive de manliga könsorganen anläggs nämligen innan fostrets kön bestämts.
Rudimentära muskler
Musculus palmaris longus är en muskel i underarmen, se figuren överst på denna sida. Den bidrar tillsammans med andra muskler till att böja handen så att handflatan närmar sig underarmen, till att kupa handflatan och eventuellt till att röra tummen. Men den saknas, i den ena eller båda armarna, hos rätt många människor. Dessutom har den ett mycket variabelt utseende hos människan. Ett par studier tyder på att att människor som saknar den inte har sämre handfunktion än andra. Möjligen kan den bidra till ett bättre handgrepp vid vissa sporter, vid vilka du griper om ett cylindriskt föremål, till exempel tennis, ishockey och rodd. Förmodligen är den viktigare hos djur, som går med frambenen. Hos människan är den sannolikt ett rudiment. Det finns flera andra muskler hos människan som har liten eller ingen betydelse. Några av dem är klart rudimentära.

Muskler som rör på ytteröronen
De flesta däggdjur kan röra på ytteröronen och rikta dem mot en ljudkälla för att höra bättre. Människan har kvar de muskler som rör på ytteröronen. Men det är bara några av oss som viljemässigt kan röra på öronen. Och vi öronviftare kan inte rikta öronen mot en ljudkälla. Alla människor har dock en reflex som gör att öronen rycker till något lite vid plötsliga ljud. Musklerna som manövrerar ytteröronen kan betraktas som rudiment.
Svansbenet ("svanskotan")
Svansen förlorades hos våra föregångare bland människoaporna. Men vi härstammar från apor med svans. Svansbenet (coccyx) hos människan består av sammansmälta ryggkotor och är en rest av den förlorade svansen. Detta ben fungerar bland annat som fäste för en muskel och ett ledband (ligament). Dessutom passerar en ryggmärgsnerv (nervus coccygeus) ut ur det. Den innehåller känselfibrer från en del av mellangården. Svansbenet har alltså vissa funktioner. Men muskeln och ligamentet fäster också på korsbenet (sacrum). Och antalet ryggkotor som ingår i svansbenet kan variera mellan tre och fem. Svansbenet kan betraktas som ett rudiment, eftersom det har liten eller ingen funktion.

Vomeronasalorganet i näsan
Vomeronasalorganet är ett särskilt luktorgan som reagerar på så kallade feromoner. Feromoner påverkar andra individer inom arten. Hos de flesta däggdjur har feromoner stor betydelse, bland annat vid fortplantningen. Hos människan är organet parigt och beläget i mellanväggen mellan de två näshålorna. Det är emellertid inte fullt utvecklat hos alla individer. Dessutom är det inte innerverat och generna för dess mottagare av doftämnen (luktreceptorerna) har förlorat sin funktion. Vomeronasalorganet kan således betraktas som ett rudiment hos människan. Men vi reagerar kanske ändå på feromoner.
Hårsäckar och gåshud
Människor är ju nästan hårlösa apor, men vi har kvar hårsäckar på hela kroppen, utom handflatorna och fotsulorna. De flesta hårsäckarna innehåller mycket små, ofta osynliga hår. Vi kan dock fortfarande resa de små håren med hjälp av mycket små hårresarmuskler. Då får vi gåshud. Forskarna har förmodat att gåshud, liksom rodnad i ansiktet, skulle fungera som en social signal, men det verkar inte vara sannolikt. De flesta av våra hårsäckar kan sannolikt betraktas som rudiment.

På bilden syns också den hålighet inuti tänderna som innehåller pulpan. Du ser också att tändernas kronor är täckta av emalj, kroppens hårdaste vävnad. Röntgenbilder är negativ. De svartaste områdena har således mottagit mest strålning. Emaljen är så tät att den absorberar nästan all röntgenstrålning. Eftersom så lite strålning passerar genom emaljen, svärtas inte filmen bakom den och den framstår som helt vit.
Visdomständerna
Under människans evolution har käkarna förkortats. Kindtänderna har också minskat i storlek. Detta är sannolikt en anpassning till en mera lättuggad föda, särskilt efter det att vi började tillaga maten med hjälp av eld. Trots att tänderna blivit mindre, har de kortare käkarna inneburit att det finns ont om plats för de bakersta kindtänderna, visdomständerna. De är ju de sista tänder som bryter fram. Ofta ställer platsbristen till med problem, som tandläkaren måste åtgärda. Ofta har de också en onormal form. Den som skriver dessa rader hade en gång en visdomstand med en J-formad rot, något som gjorde utdragandet till en mycket intressant upplevelse. Cirka 25 procent av alla människor saknar faktiskt minst en visdomstand. Det är troligt att visdomständerna är på väg att sorteras ut genom naturligt urval och att de är rudiment. Enligt en alternativ modell är visdomständernas ringa storlek betingad av embryologiska mekanismer och en direkt följd av att alla människans tänder är mindre än våra utdöda släktingars. Men denna modell förklarar inte de problem som visdomständerna ger upphov till och det faktum att de ofta saknas.
Referenser
- B. D'Aniello et al.: The vomeronasal organ: a neglected organ (Frontiers in Neuroanatomy 11:70, 2017).
- S. Benning, C. J. Patrick, och A. R. Lang: Emotional modulation of the post-auricular reflex (Psychophysiology 41:426-432, 2004).
- J. M. Bermúdez de Castro och M. E. Nicolas: Posterior dental size reduction in hominids: the Atapuerca evidence (American Journal of Physical Anthropology 96:335-356, 1995).
- R. R. Bollinger et al.: Biofilms in the large bowel suggest an apparent function of the human vermiform appendix (Journal of Theoretical Biology 249:826-831, 2007).
- S. G. Byars, S. C. Stearns, och J. J. Boomsma: Association of long-term risk of respiratory, allergic, and Infectious diseases with removal of adenoids and tonsils in childhood (JAMA Otolaryngology-Head & Neck Surgery 144:594-603, 2018).
- Jared Diamond: Father's Milk (Discover, 1 februari 1995)
- R. L. Drake, A. W. Vogl, och A. W. M. Mitchell: Gray's Anatomy for students (2nd ed, Churchill Livingstone, 2010).
- R. Elsaesser och Jacques Paysan: The sense of smell, its signalling pathways, and the dichotomy of cilia and microvilli in olfactory sensory cells (BMC Neuroscience 8 Suppl 3:S1, 2007).
- A. R. Evans et al.: A simple rule governs the evolution and development of hominin tooth size (Nature 530:477-480, 2016).
- C. Fowlie, C. Fuller, och M. K. Pratten: Assessment of the presence/absence of the palmaris longus muscle in different sports, and elite and non-elite sport populations (Physiotherapy 98:138-142, 2012).
- D. Larhammar och G. Roomans: Rudimentära organ (Vetenskap och Folkbildning, hämtad 17 oktober 2017).
- A. Pask: The reproductive system (In: D. Wilhelm, P. Bernard [eds] Non-coding RNA and the Reproductive System. Advances in Experimental Medicine and Biology, vol 886. Springer, 2016).
- S. J. Sebastin et al.: Does the absence of the palmaris longus affect grip and pinch strength? (Journal of Hand Surgery 30B:406-408, 2005).
- L. Spinney: Vestigial organs: remnants of evolution (New Scientist, 14 May 2008).
- H. F. Smith et al.: Multiple independent appearances of the cecal appendix in mammalian evolution and an investigation of related ecological and anatomical factors (Comptes Rendus Palevol 12:339-354, 2013).

Svansen är en kroppsdel som finns hos nästan alla däggdjur. Men några av dem, till exempel människan, har förlorat sin svans. Vilka funktioner har svansen? Varför har människan förlorat den?
Kaudalregion, svans och stjärt hos ryggradsdjur
Den kroppsregion som ligger bakom kroppshåligheten hos ryggradsdjur kallas kaudalregion. Det finns inga inälvor i kaudalregionen, men ryggraden fortsätter ut i den. På engelska kallar vi kaudalregionen för "tail" hos alla ryggradsdjur. På svenska däremot brukar vi använda ordet "stjärt" för en successivt avsmalnande kaudalregion som inte är väl avgränsad från kroppen och ordet "svans" för en kaudalregion som är smal och klart avgränsad. Däggdjur har således svans, medan fiskar, salamandrar och de flesta kräldjur har stjärt. Språkbruket är dock inte konsekvent. Flera pungdjur, till exempel kängurur, har en successivt avsmalnande kaudalregion, men den kallas ändå svans.

Fåglar har en mycket förkortad kaudalregion, men de fjädrar som sitter på den brukar kallas stjärt. Grodor och paddor har varken stjärt eller svans. En del däggdjur saknar helt eller i det närmaste svans, till exempel människoapor och gibbonapor, marsvinet och kapybaran, klippdassar samt koalan. Hos en del av dessa djur syns den rudimentära svansen på kroppsytan, hos andra inte. Andra däggdjur har en mycket kort svans, till exempel svin och björnar.
Kaudalregionens ursprungliga funktion är att hjälpa djuret att ta sig fram vid simning. Många fiskar simmar genom att en muskeldriven vågrörelse går bakåt längs med kroppen och pressar fisken framåt. Ormar kan röra sig på ett likartat sätt, fast på land. Andra fiskar simmar genom att slå stjärten fram och tillbaka i sidled.

Däggdjurens svans
Hos däggdjur hjälper svansen inte direkt till när djuret rör sig. Däggdjuren använder förstås benen när de rör sig. Men deras svans kan ändå vara viktig för rörelseförmågan. Ekorrar och många apor använder svansen som balansorgan och styrorgan, när de klättrar och hoppar i träden, kängurur när de hoppar och många däggdjur, till exempel geparden och andra kattdjur, när de springer. De dinosaurier som rörde sig på två ben använde förmodligen stjärten på samma sätt. När vissa kängurur rör sig långsamt, använder de svansen som ett femte ben, så kallad "femfotagång". Femfotagång innebär att kängurun först sätter ner bakbenen och sedan samtidigt framfötterna och svansen, medan bakfötterna flyttas framåt. Känguruns svans är alltså ett femte "ben", framfötterna är nämligen ganska små och behöver hjälp att hålla kroppen uppe. Myrkottar använder också svansen som stöd. När de reser sig upp stöder de sig på bakbenen och svansen.

Gripsvans, ett "femte ben"
Flera däggdjur har en gripsvans som används som ett femte ben eller en femte hand när djuren klättrar i träden. Till dessa djur hör binturongen (ett rovdjur i Sydostasien), flera myrsloksarter (som lever i Sydamerika), flera sydamerikanska apor (bland annat spindelaporna), den likaledes sydamerikanska veckelbjörnen (släkt med tvättbjörnen), de sydamerikanska gripsvanspiggsvinen, den nordamerikanska pungråttan (opossumen) och andra pungdjur samt vissa myrkottar (som finns i Afrika och södra Asien). Det finns ett svenskt djur med gripsvans, den lilla dvärgmusen (Micromys minutus).
Märkligt nog finns det många trädlevande djur med gripsvans i Amerikas regnskogar, men inga som glidflyger. I Asiens och Afrikas regnskogar finns det många trädlevande djur som glidflyger, men färre med gripsvans. En spekulation är att detta beror på att de amerikanska skogarna är tätare, men det finns inga bevis.
Gripsvans finns också hos kameleonterna bland kräldjuren.

Andra funktioner hos svansar
Svansen kan också ha andra funktioner. Den kan användas för sociala signaler. En hund kan vifta på svansen eller sticka den mellan benen. Till och med människor vet vad det betyder. Kor och hästar viftar förstås bort flugor med svansen. När flodhästen avger sin avföring, viftar den energiskt med svansen. Då sprids avföringen vida omkring, förmodligen för att markera revir. Piggsvinets taggiga svans används till försvar. Några utdöda amerikanska jättebältor (glyptodonter) använde sannolikt sin svans som en klubba, när de försvarade sig. Hos en del av dem var svansklubban spikförsedd. Hos valarna har svansen åter blivit det den en gång var hos fiskarna, det vill säga en stjärt med fena som användes till att simma med.
När och varför förlorade människans förfäder svansen?
Varför har människan ingen svans? Klart är att svansförlusten skedde långt innan det fanns människor, troligen för 15-25 miljoner år sedan hos en gemensam förfader till människoaporna och gibbonerna. Detta kan ha att att göra med att gibboner och människoapor utvecklat en ny metod att klättra i träden. Hos dessa apor spelar armarna en viktig roll. De kan svinga sig fram i träden med hjälp av armarna. De kan också gå på bakbenen längs med vågräta grenar balanserande med hjälp av armarna. Om du klättrar på detta vis har du ingen användning för svansen. Den kan till och med vara i vägen. Intressant nog började således våra förfäder sannolikt att gå på två ben redan när de levde i träden. Än i dag kan du se detta beteende hos gibboner och orangutanger, apor som tillbringar nästan all sin tid uppe i träden.
Människan och hennes svanslösa släktingar har dock fortfarande kvar ett fåtal sammanväxta svanskotor inne i koppen. De bildar svansbenet (coccyx) som sannolikt är ett så kallat rudiment. Människofoster kan utveckla en svans, men detta sker mycket sällan. Och den kan lätt opereras bort efter födseln.
Fördjupning
Var dinosaurierna och fisködlorna växelvarma (kallblodiga) eller jämnvarma (varmblodiga)? Läs om dinosauriernas kroppstemperatur under rubriken Att hålla sig varm när det är kallt, och kall när det är varmt – Om djurens kroppstemperaturreglering.
Referenser
- R. E. Blanco, W. W. Jones, och A. Rinderknecht: The sweet spot of a biological hammer: the centre of percussion of glyptodont (Mammalia: Xenarthra) tail clubs (Proceedings of the Royal Society B 276:3971-3978, 2009).
- Encyclopedia of life (hämtad 10 maj 2017).
- G. C. Hickman: The mammalian tail: a review of functions (Mammal Review 9:143-157, 1979).
- A. Horvath: Why don’t humans have tails? (Pursuit, University of Melbourne, uppdaterad 5 februari 2016).
- M. Hutchins et al.: Grzimek's animal life encyclopedia (2nd ed, Gale, 2003).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- Lars Råberg et al.: The origin of Swedish and Norwegian populations of the Eurasian harvest mouse (Micromys minutus) (Acta Theriologica 58:101-104, 2013).

Nästan alla däggdjur har märkligt nog sju halskotor. Giraffens hals är förlängd, inte genom fler kotor, utan genom att varje kota är högre. Valarna har förlorat halsen som en anpassning till vattenliv, men har fortfarande sju förkortade halskotor, dock ofta sammanvuxna. Fåglar har ett mycket varierande antal halskotor.
Däggdjurens halskotor
De enda däggdjur som inte har sju halskotor är manaterna och sengångarna. Manaterna (släktet Trichechus) tillhör sjökorna, en grupp stora vattenlevande däggdjur, som varken är valar eller sälar. De har vanligen sex halskotor. Sengångarna lever i de tropiska delarna av Amerika och hänger upp och ner i träden med hjälp av sina förlängda klor. Enligt en vederhäftig källa uppges de tretåiga sengångarna (släktet Bradypus) har 8–10 halskotor, de tvåtåiga (släktet Choloepus) 5–6 hos en art och 7 hos en annan. I litteraturen hittar du dock andra uppgifter om sengångarnas halskotsantal. Klart är emellertid att båda släktena kan ha ett halskotsantal som avviker från 7. De tretåiga sengångarna uppges kunna vrida sitt huvud upp emot tre kvarts varv (cirka 270 grader). Denna exceptionella rörlighet möjliggörs sannolikt av de övertaliga halskotorna. De tvåtåiga sengångarna uppges inte kunna vrida sitt huvud lika mycket.

Varför sju halskotor?
Varför har nästan alla däggdjur sju halskotor? Det är långt ifrån klarlagt, men det finns intressanta förslag. Halskotorna skiljer sig från bröstkotorna nedanför dem genom att de inte är anslutna till revben. Revben vid den nedersta halskotan förekommer som en ovanlig missbildning hos flera däggdjur, även hos människan. Detta innebär att denna kota förvandlats till en bröstkota. Missbildningen är kopplad till flera olika problem. Till dem hör en ökad risk att drabbas av cancer under de tidiga barnaåren. Det extra revbenet kan även utöva tryck på de nerver och den artär som försörjer armen. Detta kan ibland leda till att armen förtvinar. Möss med ett onormalt antal halskotor uppvisar en ökad cancerrisk. Allt detta borde leda till att individer med ett onormalt antal halskotor är mindre livsdugliga. Då borde också de genmutationer, som orsakar missbildningen, sorteras bort under evolutionen genom naturligt urval. En spekulativ förklaring till att manaterna och sengångarna kunnat utveckla ett avvikande halskotsantal är att dessa djur, på grund av sin låga energiomsättning, löper liten risk att drabbas av cancer.
Orsaken till halskoteabnormiteterna med de åtföljande negativa effekterna tros vara att en eller flera av däggdjurens Hox-gener inte fungerar normalt, något som stöds av molekylärbiologiska studier. Hox-generna styr embryots utveckling längs med kroppens längdaxel och därmed också anläggningen och utvecklingen av kotor och revben.
Orsakerna till att manaterna och sengångarna har ett avvikande antal halskotor är inte klarlagda. De övertaliga halskotorna hos de tretåiga sengångarna skulle kunna tolkas så att gränsen mellan halsryggraden och bröstryggraden förskjutits bakåt under fosterutvecklingen. En alternativ möjlighet är att denna gräns inte förändrats. I stället har anlagen för vävnaderna vid sidan av ryggraden, det vill säga muskler och revben, förskjutits. Dessa anlag har i så fall flyttats bakåt hos tretåiga sengångare, så att 1–3 bröstkotor förlorat revben, och framåt hos tvåtåiga sengångare, så att 1–2 halskotor har fått revben. Även andra hypoteser har föreslagits.

Fåglarnas halskotor
Fåglar har ett mycket varierande antal halskotor. De restriktioner som finns hos däggdjur finns uppenbarligen inte hos dem. Det största antalet, 25 stycken, anges för svanar. När det gäller det minsta antalet anger en källa nio hos en del små fåglar, medan en annan anger elva hos en del papegojor. Förvånande nog nämndes strutsarna ingenstans, så Anders gick till zoologiska samlingarna vid Biologiska museet och tittade. Men strutsen (som finns i Afrika), emun och kasuarerna (stora strutslika fåglar i Australien) samt nanduerna (stora strutslika fåglar i Sydamerika) hade alla betydligt färre halskotor än svanarna.
De översta halskotorna: atlas och axis
Hos både däggdjur och fåglar är den översta halskotan (atlas) och den näst översta (axis) speciellt utformade. Därmed blir huvudet mera rörligt. När vi rör huvudet framåt och bakåt (nickar) använder vi lederna mellan kraniet och atlas. När vi snurrar på huvudet, som då vi säger nej, använder vi lederna mellan atlas och axis. Axis har en tapp som sticker upp inuti atlas. Rotationen sker kring denna tapp. När vi vaggar på huvudet i sidled, använder vi både lederna mellan kraniet och atlas och de mellan atlas och axis. Dessutom sker böjningar mellan de andra halskotorna när vi rör på huvudet. Hos valar, som ju inte rör på huvudet, kan atlas och axis eller till och med alla halskotorna vara sammanväxta.

När vi nickar rör sig huvudet runt en rotationsaxel som går ungefär mellan de båda hörselgångarnas mynningar. Hos vuxna ligger huvudets tyngdpunkt framför denna axel. Det är därför som huvudet faller framåt, när du sitter i en stol och somnar. Hos spädbarn är ansiktet mindre utvecklat, så huvudets tyngdpunkt ligger bakom rotationsaxeln. Det är därför som huvudet lätt faller bakåt hos spädbarn och måste stödjas när du håller barnet.
Atlaskotan har fått sitt namn efter den jätte i den grekiska mytologin som bar hela världen på sina axlar och därvid förstås måste luta huvudet framåt. Samme jätte har gett namn åt våra kartböcker.
Fördjupning
Varifrån kommer vi? Om lansettfiskar, ryggradsdjur och människans ursprung. Läs mer under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd
Referenser
- E. A. Buchholtz och C. C. Stepien: Anatomical transformation in mammals, developmental origin of aberrant cervical anatomy in tree sloths (Evolution & Development 11:69-79, 2009).
- E. A. Buchholtz, K. L. Wayrynen, och I. W. Lin: Breaking constraint: axial patterning in Trichechus (Mammalia: Sirenia) (Evolution & Development 16:382-393, 2014).
- F. Galis: Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer (Journal of Experimental Zoology [Mol Dev Evol] 285:19-26, 1999).
- F. Galis och J. A. J. Metz: Testing the vulnerability of the phylotypic stage: on modularity and evolutionary conservation (Journal of Experimental Zoology [Mol Dev Evol] 291:195-204, 2001).
- E. Garcia Franquesa et al.: Descripción osteológica del rorcual común (Balaenoptera physalus, Linnaeus, 1758) del Museo de Ciencias Naturales de Barcelona (Arxius de Miscellània Zoològica Vol. 12, 2014).
- L. Hautier et al.: Skeletal development in sloths and the evolution of mammalian vertebral patterning (Proceedings of the National Academy of Science USA 107:18903-18908, 2010).
- M.Taylor: Long and short-necked sloths of the Cambridge University Museum of Zoology (hämtad 20 oktober 2017).
- I. Varela-Lasheras et al.: Breaking evolutionary and pleiotropic constraints in mammals, on sloths, manatees and homeotic mutations (EvoDevo 2:11, 2011).

Flera djurgrupper är segmenterade. Det innebär att djuret är uppdelat i ett mer eller mindre stort antal moduler, segment. Dessa sträcker sig från framänden till bakänden, som skivorna i ett färdiguppskuret bröd. Förvånande nog är vi människor segmenterade, precis som insekter.
Segmentens funktion och embyonala utveckling
Segmenten anläggs under embryots utveckling och är från början likadana eller nästan likadana. Under den fortsatta utvecklingen specialiseras ett mer eller mindre stort antal segment så att de får olika funktioner. Detta ger en stor evolutionär flexibilitet. När behov finns, kan ett segment ofta lätt få en förändrad funktion. Segmentering kan därför ge ett djur fantastiskt stora möjligheter att utveckla nya eller förändrade kroppsdelar, när miljön förändras.
Alla segmenterade djur har ett huvud i framänden. Det tros vara sammansatt av segment, som har förändrats så pass mycket att du inte längre kan urskilja dem. I huvudet utvecklas alltid en mun, oftast ögon och andra sinnesorgan samt en hjärna. Hos en del djur, till exempel ringmaskar och mångfotingar ("tusenfotingar"), blir nästan alla segment bakom huvudet i stort sett likadana. Hos andra djur, till exempel insekter, blir flera av dessa segment starkt specialiserade för olika funktioner. Mellankroppens tre segment är till exempel hos insekterna försedda med gångben, två av dem dessutom med vingar. Hos vissa djur, till exempel daggmaskar, är gränserna mellan segmenten tydligt markerade på djurets utsida, hos andra , till exempel fiskar, syns de inte alls där. Inälvorna i kroppshåligheterna saknar ofta tecken på segmentering efter fosterutvecklingen.
Begreppet segmentering bör begränsas till de fall då segmenten successivt växer fram ur en tillväxtzon i embryot. Det räcker alltså inte att vissa organ förekommer i flera par längs med djuret. Till blötdjuren räknas bland annat snäckor, musslor och bläckfiskar. Hos en del blötdjur finns det flera par, upp till åtta, av vissa organ, till exempel gälar, skalplattor och vissa muskler. Det har tolkats som om de äldsta blötdjuren var segmenterade. Numera anses dock detta tveksamt, bland annat därför att de upprepade strukturerna hos blötdjuren inte uppkommer successivt ur en tillväxtzon.
Vilka djur är segmenterade?
Vilka djurgrupper har då segment? Tre mycket stora djurstammar är segmenterade:
- Ringmaskar (Annelida; bland annat havsborstmaskar, daggmaskar och iglar).
- Leddjur (Arthropoda; bland annat kräftdjur, spindeldjur och insekter).
- Ryggsträngsdjur (Chordata; bland annat fiskar, kräldjur inklusive fåglar och däggdjur).
Hos vissa insekter bildas segmenten inte från en tillväxtzon, men dessa insekter härstammar med stor sannolikhet från insekter med tillväxtzon.Två mindre djurstammar, som anses ha en gemensam förfader med leddjuren, är också segmenterade: björndjur (Tardigrada) och klomaskar (Onycophora).
På videon visas klomaskarnas synnerligen märkliga jaktmetod. De sprutar ut klibbigt slem från två körtlar på huvudet. Körtlarna har ett "munstycke" och uppges ha en räckvidd på upp till 30 cm. Bytet fastnar i slemmet och kan sedan ätas upp.
Binnikemaskarnas så kallade proglottider skulle kunna betraktas som segment, men brukar inte göra det. De bildas ur en tillväxtzon, dock ej hos embryot utan hos det vuxna djuret. Binnikemaskarna är parasitiska plattmaskar inom gruppen Cestoda.
Har segment uppkommit en eller flera gånger under evolutionen?
Ryggsträngsdjur, ringmaskar och leddjur står mycket långt från varandra inom djurens släktträd. Huvudgruppen Bilateria ("tvåsidiga djur") omfattar alla bilateralsymmetriska djur. Till Bilateria räknas merparten av alla djurstammar. Undantagen är svampdjur, nässeldjur kammaneter och några mindre stammar. Bilateria indelas numera i tre överordnade grupper. Ryggsträngsdjur, ringmaskar och leddjur tillhör var sin av dessa tre grupper. Segmentering har därför sannolikt utvecklats åtminstone tre gånger under djurens evolution, ett exempel på så kallad konvergens. Om förfadern till de tre segmenterade stammarna hade varit segmenterad, skulle segmenteringen ha försvunnit hos ett 20-tal andra stammar, vilket förefaller mycket osannolikt.

Utvecklingen av segmenten i embryot styrs av ett stort antal gener. Genaktiviteten vid segmentens uppkomst uppvisar både likheter och skillnader hos de tre stora segmenterade djurgrupperna. Enligt en mycket intressant hypotes har det hos alla Bilateria ursprungligen funnits en komplicerad styrmekanism med flera samverkande gener. Denna mekanism har kunnat användas i embryot hos olika djurgrupper, men leda till uppkomsten av olika anatomiska strukturer. Möjligen har ryggsträngsdjur, ringmaskar och leddjur oberoende av varandra modifierat denna mekanism till att bilda en segmenterad kropp. Ännu så länge finns det dock inget stöd för denna hypotes, när det gäller segmentering. Men det är en ekonomisk evolutionär mekanism, som sannolikt använts i andra sammanhang.
Ryggradsdjurens segmentering
Vi kan fråga oss hur människor och andra ryggradsdjur kan vara segmenterade. Vi har ju inte några segmentgränser på kroppsytan som insekter och daggmaskar. Men ryggradens kotor och de nerver som utgår mellan kotorna återspeglar segmenteringen. Segmenteringen syns också tydligt i fiskarnas muskulatur, som är uppdelad i W- eller V-formade skivor, sedda från sidan. Under ryggradsdjurens tidiga fosterutveckling uppdelas kroppen först i en rad segment. Senare under fostrets utveckling blir det allt svårare att urskilja segmenteringen i de flesta organsystemen.

Människans segmentering
Hos människan syns segmenten inte på kroppsytan och inte heller på bålens inälvor. Men varje kota i ryggraden tillhör ett segment. Ryggmärgen är också uppdelad i segment. Från varje ryggmärgssegment löper, i anslutning till en ryggkota, två nerver till vissa bestämda delar av kroppens högra och vänstra sida. Ryggmärgssegmenten är på bilden ovan utritade som färgkodade numrerade rutor. Varje ruta svarar mot en kota och de två ryggmärgsnerver som hör till den. Segmenten är: 7 halssegment (cervikalsegment), C2–C8; 12 bröstsegment (thorakalsegment), T1–T12; 5 ländsegment (lumbarsegment), L1–L5; 5 korssegment (sakralsegment), S1–S5. De fem korskotorna är sammanvuxna till ett korsben (sacrum), som ingår i bäckengördeln. Svansbenet (coccyx) består av ett fåtal sammansmälta ryggkotor och är en rest av vår förlorade svans. En par ryggmärgsnerver löper ut genom svansbenet.
Bilden ovan visar också huden uppdelad i färgkodade dermatomer, till vänster framifrån och till höger bakifrån. En dermatom är det hudområde som via en ryggmärgsnerv leder känselinformation till ett visst segment i ryggmärgen. Gränserna mellan dermatomerna är på bilden ungefärliga. Närliggande dermatomer kan dessutom överlappa varandra.
Armar och ben uppkom först när ryggradsdjuren klev upp på land och blev fyrfotadjur. Därför syns inte segmenteringen på dem. Men på bålen är dermatomernas segmentering tydlig, något som återspeglar vår härstamning från fiskar och deras förfäder.
Fördjupning
Radiär och bilateral symmetri hos djur. Tvåsidiga djur. Varför har vi höger och vänster kroppshalva?. Läs mer under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
Referenser
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2:a upplagan, 2003).
- G. K. Davis och N. H. Patel: The origin and evolution of segmentation (Trends in Biochemical Sciences Trends in Biochemical Sciences 24:M68-M72,1999).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- C. Nielsen: Animal evolution, interrelationships of the living phyla (3rd ed, Oxford University Press, 2012).
- T. Pais-de-Azevedo et al.: Recent advances in understanding vertebrate segmentation (F1000Research 2018, 7[F1000 Faculty Rev]:97 Senast uppdaterad: 23 januari 2018).
- K. H. W. J. ten Tusscher: Mechanisms and constraints shaping the evolution of body plan segmentation (European Physical Journal E 36:54).

Skulle människan teoretiskt kunna utnyttja all energi som hon får i sig via födan? Skulle människan teoretiskt kunna klara sig utan att avge urin och avföring? Svaret på båda frågorna är naturligtvis definitivt nej. Men det hela måste redas ut.
Kan vi utnyttja all energi i födan?
Vi får i oss kemisk energi via ämnen i födan, främst i form av fett, kolhydrater och proteiner. Vid ämnesomsättningen förvandlas denna energi till en annan kemisk energiform, ämnet ATP, som är kroppens "bränsle". Samtidigt bildas vatten, koldioxid samt, vid proteinnedbrytning, även urinämne. Koldioxiden avges via lungorna, urinämnet via njurarna. Energin i ATP omsätts sedan och används vid en mängd olika processer i kroppen, till exempel muskelrörelser, blodcirkulation och nervimpulser.
Men de processer som sker i kroppen kan teoretiskt omöjligt ha en verkningsgrad på 100 procent. Därvid skiljer vi oss inte från maskiner, som bilmotorer. En del av den omsatta energin blir alltid spillvärme, precis som i en motor. Vi kan således inte omvandla all energi i födan till för oss användbar energi. Några exempel. Spillvärme uppkommer när ATP bildas under ämnesomsättningen. ATP som musklerna förbrukar kan inte omsättas helt och hållet till mekanisk energi, det vill säga muskelsammandragningar. En del blir spillvärme. Blodtrycket som hjärtat skapar förloras kontinuerligt som spillvärme på grund av friktion i blodet.

Energirik näring försvinner med avföringen
Till detta kommer att vi inte kan absorbera alla energirika molekyler från födan till blodet. En del molekyler försvinner ut med avföringen, även i viss mån sådana som vi kan bryta ner och ta upp i tarmen. Dessutom finns det molekyler i födan, som vi själva inte kan bryta ner, till exempel cellulosa, pektin och hornämnen. En del av dessa molekyler bryts i större eller mindre grad ner av grovtarmens mikroorganismer och nedbrytningsprodukterna absorberas i viss mån i grovtarmen. Men huvudelen av dessa molekyler försvinner ut med avföringen.
Avföringen består faktiskt till mycken liten del av utsöndrade ämnen som vi måste göra oss av med, alltså "slaggprodukter" eller med ett bättre ord slutprodukter. Vissa "slaggprodukter" har nämligen viktiga funktioner i kroppen, men vi måste ändå göra oss av med överskottet.
Till de få slutprodukter som lämnar kroppen med avföringen hör bland annat vissa främmande substanser (i princip gifter) och nedbrytningsprodukter av gallsalter. Men denna utsöndring är nödvändig. Avföringen består till största delen av ämnen från födan (till exempel cellulosa) som vi och våra tarmbakterier inte eller bara delvis kan bryta ner, av vatten, av tarmbakterier samt av cellrester från tarmens insida. Vi måste avge avföring. Om vi inte skulle göra det skulle mer och mer avföring ständigt ansamlas i mag-tarmkanalen. Vad som händer sedan vet jag inte, men jag kan tänka mig att tarmen till slut spricker. Möjligen dör vi dessförinnan av förgiftning. Vi skulle möjligen bli gulfärgade och förgiftade av gallämnena, precis som vid många leversjukdomar.

Njuren är inget sopnedkast
Urinen innehåller många slutprodukter. Den största mängden är urinämne (urea) som bildas vid nedbrytning av proteinernas aminosyror. Om du inte urinerar tillräckligt mycket, ansamlas urinämne i kroppen, något som till slut leder till döden. Ansamling av urinämne sker när njurarnas funktion försämras vid njursjukdom. I svåra fall måste du då rena blodet med dialys i en "konstgjord njure" utanför kroppen eller operera in en frisk njure från en donator i kroppen. Njurarna avger också merparten av de gifter vi alltid får i oss via födan, vilket naturligtvis är livsnödvändigt.
Men njurarna avger inte bara slutprodukter. De är långt mer än bara kroppens sopnedkast. De avger också nödvändiga ämnen, som vi har för höga halter av i kroppen. Har vi däremot för låga halter av sådana ämnen avger njurarna ämnena i mindre ängder. Njurarna reglerar således mycket finstämt kroppsvätskornas sammansättning på en mängd olika sätt. Detta gäller bland annat mängder och halter av vatten, natriumjoner, kloridjoner, kalciumjoner, vätejoner, vätekarbonatjoner, flera andra oorganiska joner samt flera organiska ämnen. Om vi inte bildade och avgav urin skulle vi mycket snabbt bli förgiftade, till exempel av vatten och salt som vi får i oss.
Referenser
- Anonymous: Lloyd's Bank turd from York (BBC, hämtad 26 mars 2019).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (4th ed, Sinauer, 2018).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).

Ryggradsdjurens matspjälkningssystem, mag-tarmkanalen, är en fantastisk apparat. Matspjälkningsapparaten är i princip ett långt rör, vars olika delar har olika funktioner. Den arbetar som ett löpande band, längs med vilket födan successivt transporteras och behandlas, på olika sätt i olika avsnitt. Denna grundplan är dessutom fantastiskt anpassningsbar. De olika delarna är på flera sätt anpassade till djurens föda.
Olika djurgrupper kan ha radikalt olika diet. Växtätare kan bland annat äta frukter, frön, blad eller gräs. Köttätare kan äta andra ryggradsdjur, till exempel däggdjur, fåglar eller fiskar. De kan också äta ryggradslösa djur, till exempel insekter och djurplankton. De kan fånga sitt byte själva eller äta as, alltså kadaver av döda djur.
Köttätare, växtätare och allätare
Ordet kött användes huvudsakligen om muskler hos däggdjur, fåglar och kanske fiskar. Med det problematiska ordet köttätare (karnivorer) avser jag däremot här alla djur som äter animalisk föda, inklusive de som äter insekter och andra ryggradslösa djur. Växtätare (herbivorer) äter förstås oftast växter, men ofta också alger, svampar och lavar. Allätare (omnivorer) har en blandad animalisk och vegetabilisk diet. Men det finns mycket få rena köttätare och växtätare bland däggdjuren och fåglarna. De flesta köttätare äter även en del vegetabilisk föda, de flesta växtätare en del animalisk föda. Nämnas här bör också att matspjälkning är att föredra framför "matsmältning". Födan bryts ju ner, den smälter inte som snön i vårsolen.

Mag-tarmkanalens byggnad och funktioner
Med mycket få undantag, återfinns alla bildens organ hos alla andra däggdjur. Några däggdjur saknar dock tänder och många blindtarmens maskformiga bihang. Alla fåglar saknar tänder och gom, många av dem blindtarm. Hos fåglar är en del av matstrupen utvidgad till en kräva, i vilken föda kan lagras.
Viktiga funktioner hos mag-tarmkanalen är bland annat födointag, sväljning, transport av innehållet längs med kanalen, mekanisk nedbrytning av födan, kemisk nedbrytning av födan, upptag av näringsämnen till kroppen och defekation ("bajsning").
Översikt av mag-tarmkanalens delar

Födan tas in i munhålan, i vilken sex större och flera mindre spottkörtlar mynnar. I munhålan tuggas födan och förs av tungan till svalget (pharynx). Sedan transporteras den genom matstrupen (esophagus) till magsäcken (ventriculus) och därifrån till tolvfingertarmen (duodenum), som är tunntarmens första del.
I tolvfingertarmen mynnar leverns (hepar) och bukspottkörtelns (pancreas) utförsgångar. Bukspottskörtelns huvudgång mynnar gemensamt med leverns enda gång. Gallblåsan, som lagrar galla, mynnar i leverns utförsgång. Tolvfingertarmen övergår i tunntarmens andra och tredje avsnitt (jejunum och ileum). Tunntarmen är flera meter lång. Den mynnar nedtill på bukens högra sida (se pilen) i tjocktarmen. I denna mynning finns en klaff som portionsvis tömmer ut tunntarmsinnehållet i tjocktarmen.
Tjocktarmen består av fyra delar: blindtarm (cecum), grovtarm (colon), ändtarm (rectum) och analkanal. Blindtarmen bildar en säck nedanför tunntarmens mynning. Den är försedd med ett maskformigt bihang (appendix vermiformis).
Grovtarmen består av fyra delar: en uppåtgående, en tvärgående, en nedåtgående och en S-formad. Ändtarmen avslutas med en säckformad utvidgning, ampullen, som töms genom analkanalen.
De översta luftvägarna utgörs av de båda näshålorna, svalget (pharynx), struphuvudet (larynx) och luftstrupen (trachea). Naturligtvis kan vi också andas genom munnen. Näshålorna avgränsas nedtill från munhålan av gommen, som består av en främre benförstärkt del, den hårda gommen, och en bakre del, den mjuka gommen.
Födointag och mekanisk nedbrytning
Eftersom animalisk föda oftast är lättspjälkad brukar köttätare inte tugga maten särskilt noga. Köttätande däggdjur skär ofta bara sönder maten med sina vassa kindtänder eller så krossar de den. Köttätande fåglar river sönder bytet med sin kraftiga näbb eller så sväljer de bytet helt.
Växtföda är oftast svårspjälkad. Växtätande däggdjur måste därför tugga maten väl, så att den blir lättare att angripa för de mikroorganismer, som hjälper dem med matspjälkningen. Vid idissling och så kallad koprofagi (förtäring av avföring) passerar födan växtätarnas mag-tarmkanal två gånger. Växtätande däggdjur har oftast kraftiga, slitstarka kindtänder, som mal sönder födan. Fåglar saknar tänder. De har emellertid en muskelmage, som mekaniskt sönderdelar födan, ofta med hjälp av nedsvalda små stenar. Deras näbbar är också på många olika sätt anpassade till deras födointag.

Fågelnäbben är ett fantastiskt redskap som kan anpassas till alla typer av föda. Exempel är rovfåglars spetsiga kroknäbb, som kan riva bytet i bitar, papegojors och finkars kraftiga näbbar, som kan krossa frön och frukter, insektsätande fåglars ofta smala näbbar, med vilka de fångar bytet i flykten, och många fiskätande fåglars långa näbbar, som kan fungera som spjut eller har vassa utskott, i vilka bytet fastnar.
Sväljning och transport av födan
Det förefaller inte vara särskilt funktionellt att födans väg korsar andningsluften. Det gör den hos både däggdjur och fåglar. Det finns emellertid skyddsmekanismer som gör att födan inte hamnar i luftvägarna. Här beskrivs däggdjurens sväljreflex. Vid sväljning förs den mjuka gommen uppåt och täpper till vägen mellan svalget och näshålorna. Dessutom täcks struphuvudets mynning av struplocket, samtidigt som röstspringan mellan stämbanden inuti struphuvudet stängs. Men dessa mekanismer fungerar inte alltid. Ibland kan föda komma upp i näshålorna eller ner i struphuvudet. Det förra är mest obehagligt. Det senare kan vara mycket farligt och leda till andningsstopp. Hos andra däggdjur än människan bildar struphuvudet en upphöjning i svalgregionen, vilket leder till att födan passerar på båda sidor om det, inte över det, som hos människan. Risken att födan hamnar i luftvägarna är då betydligt mindre. Människans nersänkta struphuvud är en konsekvens av att huvudets läge ändrades, när våra förfäder anpassade sig till att gå på två ben. Detta illustrerar ett viktigt faktum, nämligen att evolutionen leder till bra lösningar, men inte alltid till de bästa möjliga. Å andra sidan har det hävdats att vår talförmåga gynnas av att struphuvudet är nedsänkt.
Matstrupen är, åtminstone hos människan, överst och nederst försedd med två ringformiga slutmuskler som bara öppnas vid sväljning. Den nedre är belägen precis där matstrupen passerar mellangärdesmuskeln och får stöd av denna. Notera att magsäckens "övre magmun" inte är försedd med någon slutmuskel. När en tugga av tungan förs bakåt in i svalget, inleds sväljreflexen. Då öppnas matstrupens övre slutmuskel, födan transporteras genom matstrupen, den nedre slutmuskeln öppnas och födan hamnar i magsäcken.
Transport längs med magtarmkanalen sker med så kallad peristaltik. Ett avsnitt av den ringformade glatta muskulaturen i väggen snörper då åt hålrummet, så att det blir avsevärt smalare. Insnörpningen vandrar sedan framåt som en våg och skjuter hålrummets innehåll framför sig. Peristaltiska vågor löper med jämna mellanrum längs med magtarmkanalen, oftare när du har ätit. Enligt "tarmens lag" rör de sig lyckligtvis alltid i riktning från munnen och mot ändtarmsöppningen (anus). När du spyr, är det bukväggens skelettmuskler som pressar ihop magsäcken, så kräkning bryter inte mot "tarmens lag". Magsäckens nedersta del är försedd med en slutmuskel (pylorus). Den är hos människor märkligt nog normalt öppen, men stängs strax efter det att en peristaltisk våg fört magsäcksinnehållet mot tolvfingertarmens öppning. På så sätt portioneras magsäckens innehåll ut i tarmen i lagom takt. Ändtarmsöppningen har, åtminstone hos människor, två slutmuskler, en icke viljekontrollerad glatt muskel och en viljekontrollerad skelettmuskel, som spädbarn inte kan styra.
Kemisk nedbrytning och upptag av födan
Köttätande däggdjur och fåglar lever, som nämnts ovan, på en mera lättspjälkad föda än de flesta växtätare. De tenderar därför att ha en kortare, enklare utformad mag-tarmkanal än växtätare. I synnerhet tunntarmen, grovtarmen och blindtarmen brukar vara kortare än hos växtätare. Köttätare är däremot ofta utrustade med en stor magsäck som lagrar föda. Köttätare utsöndrar ofta också extra mycket koncentrerad saltsyra i magsäcken för att luckra upp bindväv och muskelvävnad samt lösa upp proteiner och skelettben från födan. Saltsyran i magsäcken dödar också bakterier i rutten föda, något som är särskilt viktigt för asätare. Notera att även köttätare, som normalt jagar sitt byte, också ofta äter as. En del köttätande däggdjur och fåglar spyr upp ospjälkbara delar av födan, till exempel hår och fjädrar, i form av spybollar.
Växtätande däggdjur är ofta försedda med en jäskammare, i vilken mikroorganismer hjälper dem att bryta ner födan. De kan vara framtarmsjäsare med en mycket stor magsäck, ofta uppdelad i två, tre, fyra eller fem delar. Vissa av magsäcksdelarna kan egentligen vara utvidgningar av matstrupens nedre del eller tunntarmens övre del. De kan också vara baktarmsjäsare med en mycket stor grovtarm och/eller en mycket stor blindtarm.
Märkligt nog känner vi bara till en fågelart som är framtarmsjäsare, nämligen den sydamerikanska hoatzin (Opisthocomus hoazin), som använder krävan som jäskammare. Många fåglar är dock baktarmsjäsare. En del av dem har flera blindtarmar.
Däggdjur och fåglar har således, i större eller mindre grad, hjälp av mikroorganismer med att bryta ner födan kemiskt. Men de bryter naturligtvis alla också ner den utan denna hjälp. I synnerhet köttätare och allätare bryter ner större delen av födan själva. Den kemiska nedbrytningen sker med hjälp av saltsyra i magsäcken, gallsalter från levern och en rad olika enzymer. Enzymer är proteiner som spjälkar starka kemiska bindningar i molekyler.
Magsäckens saltsyra löser upp födan och genom att bryta svaga bindningar mellan molekyler. Den spjälkar inte molekylerna. Gallans gallsalter spjälkar inte heller molekyler, utan fungerar som tvättmedel (detergenter). De är molekyler med en fettlöslig och en vattenlöslig ände. De fäster sig på ytan av stora fettdroppar, med den ena änden i fettet och den andra i det omgivande vattnet. Detta leder till att ytspänningen sänks i det vatten som håller ihop fettdropparna. Dropparna kan då slås sönder till mindre fettdroppar med en större total angreppsyta för det enzym som spjälkat fettet.

De matspjälkande enzymerna
De viktigaste enzymerna är av två typer. Här följer en beskrivning av några enzymer hos människan och många andra däggdjur.
Den ena typen av enzymer bildas framför allt i bukspottkörteln och verkar då lösta i den vätska som finns i tunntarmens hålrum. De står oftast för den inledande nedbrytningen av stora molekyler till mellanstora. Från bukspottkörteln kommer bland annat trypsin och kymotrypsin, som spjälkar proteiner, amylas, som spjälkar stärkelse, nukleaser som spjälkar nukleinsyror (DNA och RNA) och lipas som spjälkar fett (triglycerider). Pepsin bryter ner proteiner i magsäckens sura miljö. Salivamylas börjar bryta ner stärkelse i munhålan.
Den andra typen av enzymer är inbäddade i tunntarmens vägg, i de cellmembran som vetter mot dess hålrum. De bryter ned produkterna av de lösliga enzymernas aktivitet. Sådana enzymer är peptidaser, som spjälkar proteinfragment i mindre bitar, och maltas som spjälkar stärkelsefragment till druvsocker (glukos). Tarmväggens enzymer bryter också ner en del små molekyler från födan som inte kan absorberas, utan måste spjälkas till upptagbara produkter. Till de senare hör vanligt socker (sackaros), som av sukras spjälkas till druvsocker (glukos) och fruktsocker (fruktos), samt mjölksocker (laktos), som av laktas spjälkas till druvsocker (glukos) och galaktos. De flesta vuxna däggdjur saknar laktas.
Slutresultatet av alla dessa enzymers aktivitet är små molekyler, som kan absorberas och tas upp av kroppen. Dessutom finns det molekyler från födan, som inte behöver brytas ner, utan kan absorberas direkt. Till dem hör vatten. Absorptionen sker hos människan huvudsakligen i tunntarmen genom eller mellan de celler som finns närmast hålrummet. Vattenlösliga ämnen absorberas oftast med hjälp av proteintransportörer, som finns i de mot hålrummet vettande cellernas cellmembran. Sådan transport är oftast energikrävande. Fettlösliga ämnen kan däremot passera rakt igenom cellmembranen med icke energikrävande diffusion. Vatten tas upp med osmos igenom eller mellan cellerna. Osmos innebär att vatten passivt transporteras från en lösning med låg totalhalt av lösta ämnen till en med hög, så att skillnaderna i halt mellan de två lösningarna minskar. Det allra mesta vattnet absorberas hos människor i tunntarmen, resten i grovtarmen.
Näringsbehov och essentiella ämnen
Olika däggdjur och fåglar är evolutionärt anpassade till sin speciella kost. Det innebär också att olika arter skiljer sig åt i sin ämnesomsättning. Dessutom innebär det att de har utvecklat olika ekosystem av mikroorganismer i mag-tarmkanalen. Mikroorganismerna kan inte bara hjälpa dem att bryta ner många näringsämnen. De kan också producera så kallade essentiella ämnen åt dem. Essentiella ämnen är kemiska ämnen som ett djur behöver, men inte kan producera själv eller inte producera själv i tillräcklig mängd. De inkluderar hos människan vatten, flera aminosyror, några omättade fettsyror, ett flertal vitaminer och åtskilliga mineralämnen. Andra däggdjur och fåglar har nästan samma eller samma behov av essentiella näringsämnen.
Hos växtätande däggdjur bryter mikroorganismerna ner bland annat cellulosa till huvudsakligen korta fettsyror: myrsyra, ättiksyra, propionsyra och smörsyra. Fettsyrorna absorberas till blodet och är en mycket viktig energikälla. Utan mikroorganismer hade många växtätare inte fått i sig tillräcklig mycket energi med födan. Detta gäller särskilt de växtätare som äter gräs och blad, som är energifattig och svårnedbrytbar föda, Frukter och frön innehåller däremot betydligt högre halter av energirika ämnen och är lättare att bryta ner. Djur som äter animalisk föda har inte samma problem med att bryta ner födan. De kan bryta ner alla viktiga stora molekyler i födan med hjälp av egenproducerade enzymer.
Växtätarna tillgodogör sig också vitaminer och essentiella aminosyror som mikroorganismerna producerat. Växters proteiner har en annorlunda aminosyrasammansättning än djurs. För djuren essentiella aminosyror saknas därför ofta i växter, olika aminosyror i olika växter. Djur som äter animalisk föda har inte detta problem, eftersom födan har ungefär samma aminosyrasammansättning som de själva.

Några däggdjur och fåglar skiljer sig från de andra i sitt behov av ett fåtal essentiella ämnen. Ett exempel är vitamin C. Vitamin C förkommer i mycket låga halter i nästan all animalisk föda. Men de flesta däggdjur och fåglar kan bilda vitaminet själva. Hos bland annat människan är vitamin C däremot ett essentiellt ämne.
Mycket få däggdjursarter äter enbart vegetabilisk föda eller enbart animalisk föda. Växtätare äter ofta små mängder animalisk föda, till exempel insekter. Rovdjur och asätare äter ofta små mängder vegetabilisk föda. De senare kan också, precis som växtätare, tillgodogöra sig vitaminer som deras mikroorganismer producerat. Dessa tillskott i kosten kan tillföra djuren olika essentiella ämnen. Många däggdjur och fåglar äter också jord, så kallad geofagi. Funktionen med detta beteende kan vara tillförsel av essentiella mineralämnen. Mineralämnen är metalljoner. Det finns naturligtvis inga organismer, som kan bilda sådana joner. Alla organismer är beroende av att få dem i sig utifrån.
Synpunkter på människans näringsbehov
Människor kan inte leva på ren köttdiet. Detta leder förr eller senare till brist på essentiella näringsämnen. Den första bristsjukdom som uppkommer hos människan är skörbjugg, brist på vitamin C. I gamla tider drabbades många sjömän på långresor av skörbjugg. Först på 1700-talet kom vi underfund med att vi kunde bekämpa skörbjuggen med hjälp av växtföda, till exempel citrusfrukter.
Inuiternas traditionella diet består nästan bara av animalisk föda. Men de lärde sig att undvika bristsjukdomar genom att äta vissa typer av växtföda och vissa vävnader i jaktbytet. Å andra sidan finns det, till exempel i Indien, många människor som är vegetarianer. De kan klara sig bland annat genom att äta en varierad växtdiet och genom att ha en mycket välutvecklad flora av mikroorganismer i mag-tarmkanalen. Med ensidig växtdiet blir dock risken stor att drabbas av bristsjukdomar.
Människan har sannolikt ätit animalisk föda i hundratusentals år. Människan är således med stor sannolikhet en allätare, evolutionärt anpassad till att äta både animalisk och vegetabilisk föda, dock sannolikt en betydligt mindre andel animalisk föda, än de flesta av oss äter i dag. Notera att människan har en fantastisk förmåga att anpassa sig till en rent vegetabilisk och en nästan rent animalisk diet. Jag är inte kompetent att uttala mig om kostrekommendationer. Det är dock ganska lätt att visa vilket intag av ett visst näringsämne, som krävs för att undvika bristsjukdom. Det är däremot mycket svårare att visa vilken balans mellan olika näringsämnen som är bäst för vår hälsa och som minskar risken att drabbas av olika sjukdomar. Det är också svårt att fastställa värdet av eller risken med de olika dieter, som nu är populära. Officiella kostrekommendationer har delvis ändrats några gånger under de senaste decennierna. Ett råd som stått sig genom åren är dock att det är bra att äta en varierad föda. Men det är ju inte särskilt vägledande.
Förr trodde forskarna att de mikroorganismer som lever i människans tarmar, det så kallade mikrobiomet, inte hade någon större betydelse för vår hälsa. Under senare år har forskarna emellertid gjort en radikal omvärdering. Mikrobiomet är på många sätt nödvändigt för att vi ska kunna överleva. Störningar i dess sammansättning kan leda till sjukdomar.
Fördjupning
- Vart är vi på väg? Utvecklas människan fortfarande idag? Kan vi förutsäga hennes framtida evolution? Artikeln finns under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd
- Hur sker transporter i djurens kroppar? Diffusion och massflöde. Läs mer under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten
- Hur började fåglarna flyga? Om tvåbenta dinosaurier, fjädrar och vingar. Du hittar artikeln under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
Referenser
- K. E. Barrett, S. M. Barman, S. Boitano, och H. L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- C. Hickman, L. Roberts, A. Larson, H. l'Anson, och D. J. Eisenhour: Integrated principles of zoology (13th ed,, McGraw Hill, 2005).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).
- P. Willmer, G. Stone, och I. Johnston: Environmental physiology of animals (2nd ed, Blackwell, 2005).

Vad är det som avgör om ett foster blir hona eller hane, flicka eller pojke. Förvånande nog är skillnaderna mycket stora mellan olika djurgrupper. Hos vissa ryggradsdjur, bland dem människan, bestäms könet av genetiska skillnader mellan honfoster och hanfoster. Hos andra bestäms könet märkligt nog av miljön, vanligen av den temperatur som fostret utsätts för.
Könet är inte bestämt i tidiga embryon
Hos tidiga embryon av ryggradsdjur är könet ännu inte bestämt. Så småningom bildas ett par genitalkammar i kroppshålans bakre vägg. Dessa kammar innehåller en bark och en märg. Barken kan ge upphov till äggstockar (ovarier). Märgen kan ge upphov till testiklar. Föregångarna till äggcellerna och spermierna härstammar märkligt nog inte från fostret, utan från gulesäcken vägg, alltså det cellskikt som omger äggets gula. Därifrån vandrar dessa så kallade primordiella könsceller till genitalkammarna. Alla rygggradsdjur har en gulesäck, förvånande nog även däggdjuren. Däggdjurens gulesäck innehåller emellertid ingen gula, utom hos de äggläggande kloakdjuren.
Könsbestämning med könskromosomer
Könet kan bestämmas genetiskt, ofta av särskilda könskromosomer i arvsmassan. Hos människan och nästan alla andra däggdjur har hanar en X-kromosom och en Y-kromosom, honor två X-kromosomer. Hos honor inaktiveras dock den ena X-kromosomen, vilket gör att båda könen producerar lika mycket av denna kromosoms genprodukter. Den inaktiverade X-kromosomen, könskromatinet ("Barr body") kan iakttagas i honliga cellkärnor. Hos fåglar har däremot hanar två Z-kromosomer, honor en Z-kromosom och en W-kromosom.

Temperaturbetingad könsbestämning
Könet kan också bestämmas av miljöfaktorer, vanligen av den temperatur som det utvecklande embryot utsätts för under en kritisk period. Oftast finns det då en tröskeltemperatur. Vid temperaturer ovanför tröskeln utvecklas embryot till det ena könet, vid temperaturer under tröskeln till det andra. Hos vissa arter uppkommer hanar vid höga och honor vid låga temperaturer, hos andra hanar vid låga och honor vid höga. Åter andra arter har två tröskeltemperaturer. Då uppkommer hanar vid temperaturer mellan tröskelvärdena, honor både vid högre och vid lägre temperaturer.
Temperaturbetingad könsbestämning förekommer hos alla krokodildjur, hos de flesta sköldpaddor, hos några ödlor och ormar samt hos många fiskar. Kräldjur lägger ofta äggen i nästen. Krokodiler täcker över dem med växtdelar, havssköldpaddor gräver ner dem i sanden på havsstränder. Det temperaturspann, inom vilket både hanar och honor kan kläckas i ett näste, är mycket smalt. Om båda könen kläcks finns det förmodligen små temperaturvariationer inom nästet. Hos havssköldpaddor har forskare visat att vissa nästen bara ger upphov till honor, andra till hanar.

Andelen hanar och honor hos olika djur
Vid genetisk könsbestämning ligger könskvoten i regel nära 0,5, det vill säga 50 procent. Detta innebär att andelen hanar är ungefär 50 procent och andelen honor därmed ungefär lika stor. Hos människan föds cirka 52,5 procent pojkar och cirka 47,5 procent flickor. Orsaken till detta är oklar, vi kan bara spekulera. En möjlig orsak är att spermier med Y-kromosomer simmar något lite snabbare än de med X-kromosomer och därmed har större chans att nå ägget först. Å andra sidan är dödligheten något större hos det manliga könet vid alla åldrar efter födelsen och mäns medellivslängd kortare än kvinnors.
Det är synnerligen märkligt att könet kan bestämmas av temperaturen. Vid temperaturbetingad könsbestämning kan nämligen könskvoten variera mycket mer än vid genetisk. Vi kan tycka att det naturliga urvalet borde leda till 50 % honor och 50 % hanar. Den varierande könskvoten vid temperaturbetingad könsbestämning kan naturligtvis orsakas av väderlekens temperaturvariationer. Teoretiskt skulle detta dock kunna vara både en fördel och en nackdel. Men om detta vet vi mycket lite. Om populationen är liten skulle det kunna tänkas vara en fördel med en större andel honor. Då skulle populationen kunna växa snabbare. Det skulle vara mycket intressant att veta om krokodilhonor och sköldpaddshonor kan välja nästets läge efter den temperatur som råder där.
Forskare har befarat att den globala uppvärmningen skulle ställa till problem för djur med temperaturbetingad könsbestämning. Också här är kunskapsläget oklart. Å ena sidan har dessa djur upplevt stora klimatiska temperaturvariationer under sin evolution. Å andra sidan värms jordklotet nu upp snabbare än någonsin under livets utveckling. I en artikel från 2018 har forskare påvisat ett extremt överskott av honor från vissa äggläggningsstränder för soppsköldpaddan. De tror att detta beror på den globala uppvärmningen och befarar att endast honor kommer att kläckas på dessa stränder i framtiden.
Hur fungerar könsbestämningen hos människan och andra däggdjur?
Hos människan och andra äkta däggdjur (placentadäggdjur) finns det i de blivande könsorganen två komplexa mekanismer, varav den en leder till honlig och den andra till hanlig utveckling. Det handlar om genetiska kedjereaktioner, i vilka proteinprodukter av gener aktiverar andra gener. Mycket grovt förenklat skulle vi kunna säga att utveckling till hona är grundtillståndet och att Y-kromosomen stimulerar hanlig utveckling, samtidigt som den hämmar honlig utveckling.

På Y-kromosomen finns en överordnad gen som kallas Sry. Dess proteinprodukt binds till ett annat protein i cellkärnan hos de celler som senare ger upphov till testiklarnas så kallade Sertoliceller. Det bildade proteinkomplexet aktiverar en annan gen, Sox9. Via komplexa mekanismer leder detta till hanlig utveckling och hämning av honlig utveckling. Hos honor saknas Y-kromosomen och därmed genen Sry. Hos dem hämmas genen Sox9, i stället för att aktiveras. Detta sker med hjälp av proteinprodukten från genen Fox12 i de celler som senare ger upphov till äggstockarnas så kallade follikelceller. Via komplexa mekanismer leder detta till honlig utveckling.
Innan könet bestämts bildas i fostret två gångsystem, de Wolffska och de Müllerska gångarna. Hos hanfoster utvecklas de Wolffska gångarna till sädesledarna, sädesblåsorna och bitestiklarna, medan de Müllerska degenererar och bara finns kvar som rudiment. Hos honfoster utvecklas de Müllerska gångarna till äggledarna, livmodern och slidan, medan de Wolffska gångarna degenererar och bara finns kvar som rudiment.
Hos hanliga foster insöndras så småningom manliga könshomoner, androgener, framför allt testosteron, från testiklarna. Notera emellertid att det inte är könshormoner, androgener hos män och östrogener hos kvinnor, som är det första steget i de kedjerektioner som bestämmer könet. Det är närvaron eller frånvaron av Y-kromosomens Sry-gen, och därmed av Sry-proteinet, som är avgörande. Androgener är emellertid nödvändiga för de inre och yttre könsorganens fortsatta utveckling i hanliga foster. Östrogener är däremot inte nödvändiga för könsorganens utveckling i honliga foster.
Effekter av androgener och östrogener efter födelsen hos människor
Efter födelsen är androgener och östrogener, jämte andra hormoner, nödvändiga för pubertetsutvecklingen, menstruationscykeln och spermieproduktionen. Dessutom ger de upphov till de så kallade sekundära könskaraktärerna, till exempel skillnaderna i kroppsform och kroppsbehåring mellan kvinnor och män. Slutligen påverkar de hjärnan, vilket leder till skillnaderna i könsdrift och sexuellt beteende mellan män och kvinnor. Märkligt nog, påverkas könsdriften hos kvinnor troligen mest av androgener från binjurarna.
Hos de flesta däggdjur orsakar androgener helt klart aggressivt beteende hos hanar, särskilt när de konkurrerar om honor i samband med parningen. Även hos människan påverkar androgener och östrogener mycket sannolikt känslor och beteenden, dock på ett komplicerat sätt och i samverkan med andra genetiska faktorer och med miljöfaktorer. Många sådana beteenden är dock svåra att studera hos människan, bland annat beroende på varierande hormonhalter under loppet av kvinnors menstruationscykel och varierande hormonhalter i olika situationer. Dessutom finns det individuella skillnader. Mäns jämfört med kvinnors större benägenhet framför allt till dominansbeteende, men därmed också till aggression och våld, torde dock till stor del orsakas av androgener, även om andra faktorer bidrar. Effekten av testosteron hos män verkar till exempel vara olika hos olika personlighetstyper.
Könsbestämning hos fåglar
Hos nästa alla fåglar bestäms könet av könskromosomer. Som ovan nämnts, har hanar två Z-kromosomer, honor en Z-kromosom och en W-kromosom. Könet bestäms under fosterutvecklingen. Hur detta sker är oklart. Enligt en hypotes bestäms könet av genen DMRT1 som finns på Z-kromosomer. Denna gen kodar för ett protein, som är en transkriptionsfaktor. Transkriptionsfaktorer aktiverar eller inhiberar andra gener. Eftersom hanembryon har två DMRT1-gener får de en högre halt av DMRT1-faktorn än honor i sina så kallade odifferentierade gonader. Kanske är det denna skillnad som avgör om en odifferentierad gonad utvecklas i hanlig riktning till en testikel eller i honlig riktning till en äggstock. Slår du ut DMRT1-genen hos hanembryon får de äggstocksliknande gonader, om än inte normala äggstockar. Dessutom aktiveras enzymet aromatas, som omvandlar det hanliga könshormonet testosteron till det honliga könshormonet estradiol.

Könskromosomerna styr det grundläggande och första steget vid könsbestämningen. Deras aktivitet är oberoende av hanliga och honliga hormoner. Deras aktivitet leder emellertid till aktivering av gener, bland annat gener som gör att hanliga eller honliga könshormoner senare bildas. Hormonerna påverkar könsorganens utveckling och funktion i fostret och senare i livet. De påverkar också de sekundära könskaraktärerna hos vuxna individer. Till de senare hör hos många fåglar bland annat fjäderdräktens färger och kroppens storlek. Könshormonernas roll i fågelfostret är så vitt jag vet oklar.
Hur fungerar miljöbetingad könsbestämning?
Hos de flesta reptiler med temperaturbetingad könsbestämning avgörs könet med en helt annan mekanism. Vi vet inte var de receptorer som mäter temperaturen är belägna. Forskarna tror att de finns i genitalkammarna, men vi kan inte utesluta möjligheten, att genitalkammarna påverkas av receptorer i andra delar av kroppen via nerver. Hos amerikansk alligator har forskare dock visat att temperaturreceptorn TRPV4 mäter temperaturen i genitalkammarna. Inom ett visst temperaturintervall tycks denna receptor påverka gener som styr könsbestämningen. Märkligt nog verkar TRPV4 bara styra utvecklingen till hane, inte till hona. Det förefaller således finnas en annan mekanism, som styr den honliga utvecklingen.
Könet betäms hos reptilerna av de honliga hormonerna, östrogenerna. Deras mekanism är således radikalt annorlunda än däggdjurens. Enzymet aromatas tycks spela en central roll, se bilden överst på denna sida. Detta enzym är hastighetsbestämmande i den syntesväg, genom vilken östrogenet estradiol bildas. Om aromataset aktiveras ökar östrogensyntesen och fostret utvecklas till en hona. Om aromatasaktiviteten förblir låg, utvecklas fostret till en hane. Om aromataset hämmas utvecklas hanar även vid temperaturer, som normalt borde leda till honlig utveckling. Mekanismerna är emellertid dåligt utredda och det finns väsentliga skillnader mellan olika arter.
Vad kom först, miljöbetingad eller genetisk könsbestämning?
Det är mycket egendomligt att könet bestäms på så många olika sätt hos ryggradsdjuren genom radikalt olika varianter av genetisk och miljöbetingad könsbestämning. Möjligen kan könet till och med hos vissa arter påverkas av tillväxthastigheten. Hos en nejonögeart uppkommer ett kraftigt överskott av hanar i miljöer med dålig födotlllgång, där larverna växer långsamt. I miljöer med god födotillgång, där larverna växer snabbt, uppkommer även där ett överskott av hanar, men betydligt mindre. Detta kan mycket väl vara funktionellt, eftersom det kräver mer energi att producera ägg än att producera spermier.
Till detta kommer att det finns kräldjursarter vars kön bestäms både av könskromosomer och av temperaturen. Hos en skinkart med könskromosomer har hanarna antingen XX eller XY, honorna alltid XX. XX innebär två X-kromosomer, XY en X-kromosom och en Y-kromosom. Hos denna ödla gör låg temperatur under embryots utveckling att XX-embryon utvecklas till hanar. Hos en annan ödla med könskromosomer har honorna antingen ZZ (två Z-kromosomer) eller ZW (en Z-kromosom och en W-kromosom), men hanarna har alltid ZZ. Hos denna art gör hög temperatur under embryots utveckling att ZZ-embryon utvecklas till honor.
Vilken var då den ursprungliga formen av könsbestämning hos kräldjur, inklusive fåglar, och däggdjur, två grupper som med mycket stor sannolikhet har en gemensam förfader. Vi kan bara spekulera. Det finns argument för båda alternativen.
Referenser
- S. M. I. Alam et al.: Did lizards follow unique pathways in sex chromosome evolution? (Genes 9:239, 2018).
- J. M. Carré et al.: Exogenous testosterone rapidly increases aggressive behavior in dominant and impulsive men (Biological Psychiatry 82:249-256, 2017).
- J. M. Carré och N. A. Olmstead: Neuroscience forefront review. Social neuroendocrinology of human aggression, examining the role of competition-induced testosterone dynamics (Neuroscience 286:171-186, 2015).
- P. Celec, D. Ostatníková och J. Hodosy: On the effects of testosterone on brain behavioral functions (Frontiers in Neuroscience 9:12, 2015).
- S. M. Gartler et al.: Normal histone modifications on the inactive X chromosome in ICF and Rett syndrome cells: implications for methyl-CpG binding proteins (BMC Biology 2:21, 2004).
- G. C. Hays et al.: Population viability at extreme sex-ratio skews produced by temperaturedependent sex determination (Proceedings of the Royal Society B 284:20162576, 2017).
- M. Hines: Sex-related variation in human behavior and the brain (Trends in Cognitive Science 14: 448-456, 2010).
- M. Johnson Pokorná och Lukáš Kratochvíl: What was the ancestral sex-determining mechanism in amniote vertebrates? (Biological Reviews 91:1-12, 2016).
- N. S. Johnson, W. D. Swink, och T. O. Brenden: Field study suggests that sex determination in sea lamprey is directly influenced by larval growth rate (Proceedings of the Royal Society B 284:20170262, 2017).
- H. Merchant-Larios och V. Díaz-Hernández: Environmental sex determination mechanisms in reptiles (Sexual Development 7:95-103, 2013).
- N. J. Mitchell och F. J. Janzen: Temperature-dependent sex determination and contemporary climate change (Sexual Development 4:129–140, 2010).
- K. T. Moeller: Temperature-dependent sex determination in reptiles (Embryo Project Encyclopedia, uppdaterad 25 februari 2015).
- R. E. Norman et al.: Trait anxiety moderates the relationship between testosterone responses to competition and aggressive behavior (Adaptive Human Behavior and Physiology 1:312-324, 2015).
- D. O’Meally et al.: Are some chromosomes particularly good at sex? Insights from amniotes (Chromosome Research 20:7-19, 2012).
- K. A. Pavlov, D. A. Chistiakov och V. P. Chekhonin: Genetic determinants of aggression and impulsivity in humans (Journal of Applied Genetics 53:61–82,k 2012).
- R. Sekido och R. Lovell-Badge: Sex determination involves synergistic action of SRY and SF1 on a specific Sox9 enhancer (Nature 453:930-934, 2008).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).
- R. Yatsu et al.: TRPV4 associates environmental temperature and sex determination in the American alligator (Scientific Reports 5:18581, 2015).

Regeneration innebär att en individ kan återbilda en förlorad kroppsdel eller att en avskuren bit av en individ kan utvecklas till en hel individ. Förmågan att regenerera är olika stor inom olika djurgrupper. Vissa enkelt byggda djur kan regenerera en helt ny individ från en liten bit av kroppen. Andra djur, till exempel däggdjur och de flesta insekter, saknar helt eller nästan helt förmågan att återbilda kroppsdelar, åtminstone som vuxna.
Stamceller och deras funktion
För att regeneration ska ske krävs i de flesta fall stamceller. Stamceller är odifferentierade (omogna) celler som kan dela sig och därvid ge upphov dels till nya stamceller, dels till celler med högre differentieringsgrad. Totipotenta stamceller kan ge upphov till alla de celltyper som ett befruktat ägg kan ge upphov till under embryonalutvecklingen. Multipotenta stamceller kan ge upphov till flera celltyper, men inte alla. Unipotenta stamceller kan bara ge upphov till en celltyp, till exempel skelettmuskelceller eller nervceller. Hos däggdjur finns dessutom pluripotenta stamceller som kan ge upphov till alla celler i embryot utom cellerna i den yttre fosterhinnan. Regeneration stimuleras ofta av tillväxtfaktorer, av hormoner eller av större molekyler i den så kallade extracellulära matrixen utanför cellerna. Vid regeneration av komplicerat byggda delar av kroppen, till exempel ben och inre organ, krävs dessutom en återaktivering av de intrikata mekanismer som styrde formbildningen (morfogenesen) under embryots utveckling. Formbildning innebär att strukturer i kroppen utvecklas från en ostrukturerad vävnad, till exempel ett ben från en extremitetsknopp.
Hydror och plattmaskar kan bilda en helt ny individ ur en avskuren bit av kroppen
Hydror är nässeldjur med fantastisk regenerationsförmåga. En hydra består av ett skaft som är fäst i underlaget med en fot. Överst är skaftet försett med en krans av tentakler kring munnen. Hydror kan skäras i flera bitar, som alla kan bilda fullständiga nya individer. Ett skaft kan bilda en överdel och en överdel kan bilda ett skaft.

Plattmaskarna är en enkelt uppbyggd djurgrupp. Bland plattmaskarna är virvelmaskar (klassen Turbellaria) kända för sin regenerationsförmåga. Du kan dela en del virvelmaskar i flera bitar, som alla tillväxer och ger upphov till en ny fullständig mask, med huvud och allt. Hos en plattmaskart har forskarna i vuxna maskar identifierat totipotenta stamceller. Dessa celler kan, precis som det befruktade ägget, via celldelningar ge upphov till alla de celler som finn i en vuxen mask, inklusive könscellerna. En enda sådan cell kan rädda en mask, som utsatts för dödlig radioaktiv strålning, så att den åter blir en normal, frisk individ.
Slemmaskar (Nemertea) har också en mycket god regenerationsförmåga. En liten bit av en mask kan ge upphov till en fullständig ny individ. Rundmaskar (Nematoda) regenererar däremot dåligt. De kan i stort sett bara plugga igen ett hål i kroppsväggen.

Daggmaskar har begränsad regenerationsförmåga
Daggmaskarna tillhör gruppen fåborstmaskar inom stammen ringmaskar (Annelida). Till ringmaskarna hör dessutom iglarna och de marina havsborstmaskarna. Ringmaskarnas kropp är uppdelad i ett flertal segment. Havsborstmaskar kan vara bra på att regenerera. Hos ett par arter kan en komplett mask uppkomma ur ett enda bortskuret segment. Daggmaskar kan också regenerera så att, i vissa fall, två fullständiga maskar uppkommer om masken skärs i två bitar. Olika arter har dock olika väl utvecklad regenerationsförmåga. Ofta finns det vissa begränsningar. En framdel kan vanligen ge upphov till en fullständig mask. En bakdel kan oftast bara ge upphov till ett visst maximalt antal segment, olika antal hos olika daggmaskarter. Det innebär att en alltför kort bakdel utvecklas till en ofullständig mask. En sådan ofullständig mask kan bland annat sakna könsorgan. Den vanliga daggmasken Lumbricus terrestris är en art med begränsad regenerationsförmåga. Iglar kan inte regenerera alls.

Återväxt av förlorade armar och ben
Mer komplicerat byggda djur kan i regel inte regenerera nya individer om kroppen skärs i bitar. Däremot kan många av dessa djur regenerera förlorade extremiteter. Detta gäller för de flesta kräftdjur. Hos en del insekter kan förlorade ben växa ut igen, men bara hos djur som är i larvstadiet. Bland blötdjuren lär bläckfiskar kunna regenerera förlorade armar. Bland tagghudingarna är sjöstjärnorna välkända för att kunna regenerera förlorade armar.
Stjärtgroddjurs fantastiska regenerationsförmåga
Bland ryggradsdjuren är stjärtgroddjuren (Urodela), till vilka våra vattensalamandrar hör, kända för att ha en fantastisk regenerationsförmåga. De kan bland annat regenerera förlorade ben och till och med käkar. Grodlarver kan också regenerera förlorade ben, men det kan inte vuxna grodor. En del ödlor kan regenerera en förlorad stjärt. Stjärten bryts av på ett bestämt försvagat ställe. Sådan så kallad autotomi kan var ett sätt att slippa undan ett rovdjur.
Däggdjur har nästan helt förlorat förmågan till att regenerera kroppsdelar
Vi däggdjur kan inte regenerera förlorade ben och armar. Men vi kan regenerera en del inre organ och vissa vävnader, om det finns en bit kvar av organet eller vävnaden. Levern och njurarna är kända för detta. Om större delen av levern tas bort, kan återstoden regenerera till en lika stor ny lever, om än inte med samma form som den gamla. Om en njure opereras bort, tillväxer den andra och tar över dess funktioner. Nya skelettben kan inte bildas, men benvävnad kan regenerera vid läkning av benbrott och ny överhud kan bildas vid sårläkning. Blodets celler, blodkropparna, nybildas ständigt, liksom det innersta lagret av celler (epitelet) i mag-tarmkanalen och överhudens celler, som ger upphov till dess hornlager.

Människor, åtminstone barn, kan också regenerera den allra yttersta delen av ett finger, förutsatt att nagelns tillväxtzon (matrix) finns kvar. I denna zon finns stamceller som krävs för regenerationen. Regenerationen inkluderar bildning av ny benvävnad, vilket innebär att den kvarvarande stumpen av det yttersta skelettbenet förlängs. Kanske är detta det enda vi kvar av den förmåga att regenerera ett helt gångben som finns hos stjärtgroddjuren. En intressant spekulation är att däggdjurens förlorade regenerationsförmåga åtföljs av en minskad cancerrisk och att nettoeffekten skulle vara till fördel för djuren. Forskare har också gissat att den minskade regenerationsförmågan möjliggör en snabbare sårläkning genom ärrbildning i stället för regeneration.
Kan vi återuppväcka regenerationsförmågan, så att förlorade armar och ben återbildas?
Det är dock möjligt att vi däggdjur har kvar de mekanismer som gör att groddjur kan regenerera ett helt ben, men att de styrande generna är avstängda överallt utom i fingertopparna och kanske tåspetsarna. Denna tanke stöds av resultat från försök med möss. Några proteiner, som är nödvändiga för regeneration av deras framtåspetsar, återfinns också hos stjärtgroddjur. Dessa proteiner har hos groddjuren samma funktioner som hos mössen.
Musstammen MRL har oväntat visat sig ha fantastiska egenskaper. Den är känd för att den drabbas en viss immunsjukdom. Men dessa möss uppvisar också en anmärkningsvärd regenerationsförmåga, möjligen en följd av att de inte bildar ärr vid läkning av skador. De kan till och med regenerera hjärtmuskelvävnad. Om vi kunde åstadkomma hjärtmuskelregeneration hos människor skulle vi kanske kunna minska dödligheten i hjärt-kärlsjukdomar. En del forskare menar att vi skulle kunna återuppväcka förmågan till regeneration hos människor helt enkelt genom att blockera ärrbildning. Men vi kan inte utesluta möjligheten att cancerrisken då skulle öka.
Det bedrivs mycket forskning om regeneration, särskilt med axolotlen (Ambystoma mexicanum) och andra stjärtlösa groddjur som försöksdjur. Det yttersta målet med denna forskning är inte bara att reda ut regenerationsmekanismerna hos groddjuren, utan också att kunna återuppväcka förmågan till regeneration hos sjuka och skadade människor, men det dröjer nog innan vi är där.
Fördjupning
Hur kryper daggmaskar och plattmaskar? Artikeln finns under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
Referenser
- B. I. Balinsky: An introduction to embryology (3rd ed, Saunders, 1970).
- R. B. Borgens: Mice regrow the tips of their foretoes (Science 217:747-750. 1982).
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2:a upplagan, 2003).
- B. S. Douglas: Conservative management of guillotine amputation of the finger in children (Australian Paediatric Journal 8:86-89, 1972).
- A. Heydemann: The super super-healing MRL mouse strain (NIH Public Access, Frontiers in Biology (Beijing) 7: 522-538, 2012).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012 ).
- M. Takeo et al.: Wnt activation in nail epithelium couples nail growth to digit regeneration (Nature 499:228-234, 2013).
- C. Wilson: The regeneration game (New Scientist, 27 September 2003).
- E. Yong: How nails regenerate lost fingertips (Nature, 12 June 2013).
- A. Zeng et al.: Prospectively isolated tetraspanin+ neoblasts are adult pluripotent stem cells underlying Planaria regeneration (Cell 173:1593-1608, 2018).

Däggdjurens hornbeklädnader har i alla tider spelat en stor roll i människans föreställningsvärld. Det finns hornbeklädda människofigurer på bronsålderns svenska hällristningar. Förmodligen bär de hornförsedda hjälmar. Den keltiske guden Cernunnos bar hjorthorn på huvudet. Sibiriska schamaner har in i sen tid använt horn som huvudbeklädnad. Den mytiska enhörningen se ofta i medeltida konst och förekommer på vapensköldar. Den bär ett långt horn på pannan.
Hos däggdjuren förekommer horn på hjässan hos familjerna slidhornsdjur, hjortdjur, gaffelantiloper och giraffdjur bland de partåiga hovdjuren (ordningen Artiodactyla). Horn på nosen finns hos familjen noshörningar bland de uddatåiga hovdjuren (ordningen Perissodactyla).
Noshörningar
Noshörningarna (familjen Rhinocerotidae) har ett eller två horn som är unika på grund av sin placering på nosen. De skiljer sig också från andra horn genom att de uteslutande bildas av överhuden (epidermis) och huvudsakligen består av tätt sammanpressade hårliknande fibrer av keratiner (hornämnen). De har således ingen kärna av benvävnad.
Slidhornsdjur
Till slidhornsdjuren (familjen Bovidae) hör bland annat får, getter, myskoxar samt oxdjur inklusive bufflar. Slidhornsdjurens två horn består av en kärna av benvävnad, kvicken. Benkärnan är täckt av hud: överhud, läderhud och underliggande bindvävnad. Överhuden ger ytterst upphov till ett tjockt lager av döda celler, slidan. Slidan motsvarar hudens yttersta lager, hornlagret. Dess celler är fyllda med keratiner (hornämnen). Kvicke och slida tillväxer hela livet. Hornen fälls aldrig och är aldrig grenade. Hos större arter bär vanligen både honor och hanar horn. Hanarnas horn är dock oftast större. Hos mindre arter saknar honorna ofta horn. Det finns stora variationer i hornens form (raka, böjda eller spiraliserade), ytstruktur (släta eller räfflade) samt i deras storlek.
Gaffelbocken
Den amerikanska gaffelbocken eller gaffelantilopen (familjen Antilocapridae) har två horn med kvicke av ben och slida av hornämnen, precis som slidhornsdjuren. Till skillnad från slidhornsdjuren fäller emellertid gaffelbockarna hornets slida varje år, varefter en ny slida bildas. Både hanar och honor har horn. Hanarnas är större. Åtminstone hos äldre hanar är hornen tvågrenade med en framåtriktad tagg. Taggen består märkligt nog bara av slida, kvicken är således inte förgrenad. Gaffelbocken är den enda kvarlevande arten inom sin familj. De är känd för att vara den kanske främsta uthållighetslöparen bland däggdjuren.
Hjortdjur
Hos hjortdjuren (familjen Cervidae) bildas två horn som fälls, vanligen varje år. Nya horn växer ut från ett område på skallen som är omgivet av en benring, rosenkransen. Hornen består av benvävnad. Medan de växer ut är de täckta av hårbeklädd levande hud: överhud, läderhud och underst bindvävnad. Denna hud kallas basthud. Blodkärl från basthuden förser hornens benvävnad med de näringsämnen som behövs för dess tillväxt. När hornen växt färdigt, dör basthuden och benvävnaden blottas. Djuren gnider då av huden mot till exempel trädstammar, så kallad fejning. Hornen är hos de flesta arter förgrenade, med två eller flera grenar. Vanligen är de trädlikt grenade, cervina horn. De kan också vara så kallade skovelhorn, palmata horn. Då är "trädets" stam tillplattad, som hos älg och dovhjort. Hos älgen förekommer även cervina horn. Hos nästan alla hjortdjur förekommer horn bara hos hanarna. Hos renen har dock både hanar och honor horn. Hos det kinesiska vattenrådjuret saknar båda könen horn. Ju äldre en hane är, ju större är hans horn. Mycket gamla hanar kan dock ha horn, som blir allt mindre med åren.

Giraffdjur
Det finns bara två kvarlevande arter bland giraffdjuren (familjen Giraffidae): okapin och giraffen. Deras horn är benutväxter från skallen, som är klädda med hårförsedd hud: överhud, läderhud och bindvävnad. Giraffen utmärker sig genom att den kan ha upp till fem horn.
Hur bildas horn?
Vi vet inte mycket om hur de partåiga hovdjurens horn anläggs. Men de tycks bildas på olika sätt hos de olika grupperna, antingen som utväxter under pannbenets benhinna eller från separata benbildningscentra i underhuden. Många menar att slidhornsdjur, gaffelantiloper, hjortdjur och giraffdjur har utvecklat horn oberoende av varandra. Andra menar att alla grupperna härstammar från en hornbeklädd förfader och att hornens struktur har modifierats inom varje grupp under dess utveckling.
Sexuell selektion
Hornen hos de hornbeklädda djuren har nästan alltid en funktion vid fortplantningen. De är föremål för den evolutionära process, som kallas sexuell selektion. Hornen används ju när hanarna kämpar om att få para sig med honorna. Honorna kan också tänkas föredra hanar med stora och välväxta horn, som kan vara ett tecken på fortplantningsduglighet och en högkvalitativ genuppsättning.
Referenser
- E. B. Davis, K. A. Brakora, och A. H. Lee: Evolution of ruminant headgear: a review (Proceedings of the Royal Society B 278:2857-2865, 2011).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- R. Kuehn et al.: Molecular phylogeny of Megaloceros giganteus - the giant deer or just a giant red deer? (Zoological Science 22:1031-1044, 2005).
- J. F. Lemaître et al.: The allometry between secondary sexual traits and body size is nonlinear among cervids (Biology Letters 10:20130869).
- A. M. Lister: The evolution of the giant deer, Megaloceros giganteus (Blumenbach) (Zoological Journal of the Linnean Society 112:65-100, 1994.
- C. McDowall: The lady and the unicorn and 'millefleurs' style tapestries (The Culture Concept Circle, juni 21, 2014).
- P. Maity och S, A. Tekalur: Finite element analysis of ramming in Ovis canadensis (Journal of Biomechanical Engineering 133:021009, 2011).
- N. Solounias: Evidence from horn morphology on the phylogenetic relationships of the pronghorn (Antilocapra americana) (Journal of Mammalogy 69:140-143).

I stort sett alla färger hos människans och andra däggdjurs överhud och hår åstadkoms av stora molekyler som kallas melaniner. Ett undantag är den röda hudfärg som uppkommer när den röda färgen hos blodets hemoglobin lyser igenom svagt pigmenterad hud. Hos fåglar ger melaniner upphov till hudfärgen och de flesta fjäderfärger. Märkligt nog bildas melaninerna inte i överhuden, utan i celler som vandrar ut från det embryonala nervsystemet. Hos fåglar förekommer dessutom andra färgämnen än melaniner.
Färger orsakade av färgämnen, pigment, kallas pigmentfärger. Framför allt fåglar, men även däggdjur, kan dessutom ha så kallade strukturfärger, som inte orsakas av färgämnen, utan av optiska effekter i ofärgade strukturer. Ofta kan en pigmentfärg tillsammans med en strukturfärg ge upphov till en annan färg. Gult och blått kan till exempel ge upphov till grön färg.
Melaniner ger svarta, bruna, röda och gula färger
Melaniner är polymerer, det vill säga kedjor av små sammanbundna molekyler. De små molekylerna bildas genom kemisk modifiering av aminosyran tyrosin. I hud, hår och fjädrar hos däggdjur och fåglar finns det två typer av melaniner. Eumelaniner ger svarta, grå och bruna färger, feomelaniner gula och röda. Melaninerna bildas i pigmentceller som kallas melanocyter. Jag beskriver först hur huden färgas av melaniner hos däggdjur och fåglar. Färgningen av hår och fjädrar går till på ett likartat sätt.

Pigmentceller vandrar in i huden från neurallisten
Melaniner bildas av pigmentceller som kallas melanocyter. Melanocyterna härstammar från den så kallade neurallisten, som uppkommer när anlaget till det centrala nervsystemet bildas under embryonalutvecklingen. Detta anlag utgörs av en ränna som buktar in från ektodermet, den embryonala huden, på ryggsidan och sedan slutes till ett rör, nervröret. Neurallisten bildas i gränsområdet mellan rännan och hudektodermet. Märkligt nog vandrar sedan omogna melanocyter från neurallisten genom hela kroppen ut till överhuden.

Hur hudens hornlager bildas och får sin färg
Överhuden (epidermis) består av ett ständigt växande undre lager av levande celler, så kallade keratinocyter, och ett övre hornlager av döda keratinocyter. Längst ner i överhuden delar sig keratinocyterna. De nybildade keratinocyterna förskjuts uppåt, samtidigt som de bildar de motståndskraftiga proteiner som kallas keratiner (hornämnen). Till slut är cellerna helt fyllda av keratiner. Då dör de och bildar hornlagret. Hornlagret slits ständigt genom att de yttersta döda cellerna faller av. De avfallande cellerna ersätts emellertid ständigt underifrån av nybildade hornlagerceller. Därmed förblir hornlagret lika tjockt, dock olika tjockt på olika kroppsdelar. Hos människor är det tjockast på fotsulorna. Vid kraftigt slitage nybildas hornlagret snabbare och blir tjockare, till exempel på händerna vid långvarigt hårt arbete. Forskarna vet inte hur denna skyddsmekanism regleras.

De omogna melanocyterna delar sig även de och mognar. De är belägna i överhudens understa lager eller, hos fåglar, i den underliggande läderhuden. Där bildar de en mängd långa utskott, dendriter, som utbreder sig i överhuden. De bildar nu också melaniner. Melaninerna paketeras i en mängd membranomgivna små korn, som kallas melanosomer. Melanosomerna vandrar ut i dendriternas spetsar. Sedan sker något mycket egendomligt. Melanosomerna överföres från melanocyterna till keratinocyterna. Forskarna vet fortfarande inte hur detta går till. En hypotes är att keratinocyterna bildar inbuktningar runt dendritspetsarna och att dendritspetsarna snörps av, samtidigt som inbuktningarna sluts och bildar membranomgivna blåsor inuti keratinocyterna. Detta kallas fagoendocytos. Hur som helst, så är keratinocyterna nu färgade av melanosomkornen.
Hur hårstrån bildas och får sin färg
Hår består, precis som hornlagret, av döda keratinocyter som färgats av melanosomer. Ett hårstrå växer ut från en nedsänkning i huden, en follikel. Tillväxten sker längst ner i follikeln, där levande keratinocyter delar sig. De keratinocyter, som bildas, förskjuts uppåt när nya tillkommer. Så småningom fylls de med keratiner och dör.
Möjligen fungerade de första håren som känselorgan, precis som morrhåren hos dagens däggdjur. Fossil av kranier antyder att morrhår fanns hos förfäder till däggdjuren. Kanske utvecklades en tät värmeisolerande päls av hår först i ett senare utvecklingsskede. Å andra sidan bevaras hårstrån ytterst sällan som fossil, så pälsen kan mycket väl ha kommit först.

Hur fjädrar bildas och får sin färg
Bilden ovan visar hur en färdig konturfjäder är uppbyggd. Fjädrar bildas av överhuden (epidermis) och består precis som överhudens hornlager av döda keratinocyter. En fjäder växer ut ur en fördjupning i överhuden, en follikel. Follikeln innehåller nederst en papill, försedd med en ring av keratinocyter som delar sig. De nybildade keratinocyterna förskjuts uppåt, varvid en cylinder av keratinocyter uppkommer, förlängs och skjuter upp ut follikeln. Den består av två skikt: ytterst en skida, innerst fjäderanlaget. Skidan brister så småningom och bryts ner. I fjäderanlaget anläggs redan i papillen fjäderns skaft och strålar i form av förtjockade åsar. I cellskikten mellan de blivande strålarna utbildas bistrålarna med sina hakar. Så småningom spricker dessa cellskikt i mitten. Då blir strålarna med vidhängande bistrålar fria och kan vecklas ut till en färdig fjäder. Vid det här laget är keratinocyterna döda och fyllda med keratiner.

På papillens yta inuti fjäderfollikeln finns en ring av omogna melanocyter. Dessa delar sig hela tiden och bildar nya melanocyter, som successivt inkorporeras i fjäderanlaget och följer med uppåt när anlaget växer. Melanocyterna mognar och bildar melanosomer, som vandrar ut i dendriter. Melanosomerna överförs sedan till fjäderns keratinocyter på samma sätt som beskrivits för huden. De hamnar så småningom i den färdiga fjäderns döda keratinocyter.
Hur fjädrar blir mönstrade
Mönstrade fjädrar kan tänkas uppkomma på olika sätt. Här är några möjliga exempel. Om ringen av omogna melanocyter periodvis upphör att dela sig eller upphör att bilda melaniner, borde en tvärstrimmig fjäder bildas. Vita band på fjädern motsvarar den tidsperiod då inget melanin tillförs fjädern. Om olika melanocyter i ringen bildar olikfärgade kombinationer av melaniner, borde den färdiga fjädern få olika färger. Forskarna vet emellertid mycket lite om hur fjädrarnas färgmönster uppkommer.
Fåglar kan ha andra färgämnen än melaniner
Hos många fåglar kan andra färgämnen än melaniner byggas in i fjädrarna, Vanligast bland dem är röda, oranga eller gula karotenoider, samma typ av färgämnen som finns i morötter och som gör höstlöven gula. Mera ovanliga är porfyriner, metalloxider, psittakofulviner och sfeniskiner. Karotenoider och porfyriner tas upp via födan. Psittakofulviner finns bara hos papegojor och bildas troligen av fåglarna själva. De gula sfeniskinerna finns bara hos pingviner och är dåligt kända.

Pigmentfärger och strukturfärger
Ovan diskuterade färger är pigmentfärger. De orsakas av färgämnen, som absorberar vissa våglängder i vitt ljus och reflekterar andra. De reflekterade våglängderna når betraktarens ögon och bestämmer därmed färgen. Hos fåglar finns emellertid även så kallade strukturfärger. De orsakas inte av färgämnesmolekyler, utan av fjädrarnas uppbyggnad. Det handlar om optiska fenomen som uppkommer, när ljus träffar fjädrarna, nämligen reflektion, absorption, spridning och interferens. I många fall kan fjädrarnas färg orsakas av en kombination av en pigmentfärg och en strukturfärg. Grön färg hos undulater orsakas till exempel av en gul pigmentfärg tillsammans med en blå strukturfärg.
Strukturfärger uppkommer genom optiska effekter utan att färgämnen behövs
Reflektion innebär att ljus "studsar" tillbaka, när det träffar en yta. Absorption innebär att ljuset försvinner och omvandlas till värme, när det träffar en yta. Spridning innebär att ljus reflekteras i alla riktningar, när det träffar molekyler, mycket små partiklar eller mycket små luftblåsor.
Interferens beror på att ljus är en vågrörelse, med toppar och dalar. Se bilden nedan. Om ljus sprids eller reflekteras från två eller flera ytor som ligger på olika nivåer, mycket nära varandra och på avstånd från varandra motsvarande ljusets våglängder, kan ljus av en viss våglängd förstärkas genom interferens. Detta beror på att de två reflekterade ljusvågorna av denna våglängd hamnar i fas med varandra, så att deras toppar sammanfaller och ljuset förstärkes. Andra våglängders ljusvågor hamnar ur fas med varandra, så att deras ljus försvagas eller släcks ut. Eftersom ljusets färg bestäms av dess våglängd, kommer det resulterande ljuset att domineras av en viss färg. Du kan iakttaga interferens på en såpbubbla. Interferensen orsakas av reflektion mot de inre och yttre ytorna på såpbubblans tunna vägg. Den regnbågsliknande effekten beror på att väggen är olika tjock på olika ställen. Därför kommer olika färger förstärkas i olika väggpartier.

Iriserande strukturfärger
Iriserande strukturfärger har ett metalliskt, regnbågsliknande skimmer. Färgnyansen tenderar att skifta, när fjädern betraktas från olika vinklar. Iriserande fjädrar innehåller mycket små reflekterande plattor, som ligger olika djupt i fjäderns strålar. Avståndet mellan dem är på nanonivå, det vill säga i samma storleksordning som det synliga ljusets våglängder. Det för människan synliga ljuset upptar våglängdsområdet 390–770 nanometer (1 nanometer, förkortat nm, är en miljondels millimeter). När ljuset reflekteras mot plattor på olika djup i fjädern, kommer de reflekterade ljusvågorna inte vara i fas med varandra, se bilden ovan. Vissa våglängder kommer därför att i mer eller mindre hög grad förstärkas med hjälp av interferens, medan andra våglängder försvagas eller släcks ut. Detta resulterar i en färg som är beroende på avståndet i djupled mellan plattorna. Om betraktningsvinkeln ändras, kommer ljusets väg ner mellan plattorna bli längre eller kortare. Då kommer en annorlunda färg att förstärkas med interferens. Det är därför som iriserande fjädrar ofta är regnbågsskimrande. Välkända fåglar med iriserande fjädrar är påfåglar, kolibrier och paradisfåglar.

Icke iriserande strukturfärger
Icke iriserande strukturfärger uppkommer genom en kombination av spridning, interferens och absorption. Mekanismerna är dock inte fullständigt utredda. Blå färger är nästan alltid icke iriserande strukturfärger. Fjädrarna kan då innehålla ett yttre lager som ser "svampartat" ut på grund av en mängd mycket små luftbubblor. Ljus reflekteras mot dessa bubblor och sprids i alla riktningar. Det är främst det kortvågiga blå, violetta eller ultravioletta ljuset som sprids. Detta är i princip samma mekanism som gör att himlen blir blå. Ljus av andra våglängder passerar igenom det svampiga lagret och absorberas av ett underliggande lager med svarta melaninkorn. Färgen förstärks troligen i de flesta fall genom interferens. Vi talar då om koherent spridning. Resultatet kan då till exempel bli blå färg. Exempel är den blå färgen hos undulater och blåmesens klarblå hätta. Ett annat exempel den blå ansiktsfärgen hos hanarna av mandrillen, en babianart. Blå färger är emellertid mycket sällsynta hos däggdjur.

Enligt en undersökning från 2012 uppkommer dock de olika färgerna på ett mera komplicerat sätt än vad som beskrivits ovan. Denna studie tyder bland annat på att de gröna fjädrarna har både blå och grön strukturfärg, men att den blå färgen absorberas av färgämnen.
Vit färg
Vita fjädrar saknar helt färgämnen. De innehåller mycket små luftbubblor som i lika hög grad sprider alla färger inom det för människan synliga spektralområdet, så kallad totalreflektion. Detta gör att de ter sig vita för våra ögon. Vitt ljus är ju en kombination av alla regnbågens färger. Det är samma fenomen som gör mjölk vit, fast i mjölken sprids ljuset av små fettdroppar.
Varför är himlen blå?

Himlens färg är en strukturfärg. Luften innehåller inga färgämnen och är därför färglös. Solljuset innehåller alla regnbågens färger och uppfattas därför som vitt. Men när solljuset passerar genom atmosfären, träffar det kortvågiga blå ljuset luftens molekyler och sprids i alla riktningar, också ner mot jordytan. Därför ser himlen blå ut. Den blå färgen åstadkoms bara genom spridning av ljuset, så kallad inkoherent spridning. Den förstärks inte genom interferens som den blå färgen i fågelfjädrar. Det långvågigare gröna, gula, oranga och röda solljuset sprids inte lika mycket, utan fortsätter till största delen rakt fram, så att det inte når betraktaren. Det violetta och indigoblå solljuset är kortvågigare än det blå och sprids även det. Men dessa färger absorberas till stor del högt uppe i atmosfären. Dessutom är våra ögon mindre känsliga för dem. Därför ser vi violett och indigoblått på himlen bara i regnbågar.
Fördjupning
- Varför är så många fåglar vita? Om fjäderdräktens färger hos fåglar Artikeln finns under rubriken Förnuft, känslor och beteenden – Hur funkar hjärnan, nervsystemet, sinnen och hormoner?
- Hur började fåglarna flyga? Om tvåbenta dinosaurier, fjädrar och vingar Läs mer under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
- Hur stor kan du vara och ändå flyga? Om stora fåglar och flygödlor Du hittar artikeln under rubriken Varför är vissa djur stora medan andra är små?
Referenser
- R. E. Boissy: Melanosome transfer to and translocation in the keratinocyte (Experimental Dermatology Suppl. 2 12:5-12, 2003).
- R. E. Boissy, L. S. Trinkle, och J. J. Nordlund: Neural-tube-derived melanocyte subsets undergo commitment to their distinct lineages in culture (Cell Differentiation and Development 30:129-145, 1990).
- L. D'Alba, L. Kieffer, och M. D. Shawkey: Relative contributions of pigments and biophotonic nanostructures to natural color production: a case study in budgerigar (Melopsittacus undulatus) feathers (Journal of Experimental Biology 215:1272-1277, 2012).
- D. Dhouailly et al.: Getting to the root of scales, feather and hair: As deep as odontodes? (Experimental Dermatology 1-6, 2017).
- C. M. Eliason och M. D. Shawkey: Decreased hydrophobicity of iridescent feathers: a potential cost of shiny plumage (Journal of Experimental Biology 214:2157-2163, 2011).
- P. Gibbs: Why is the sky blue? (Original Usenet Physics FAQ, 1997).
- M. Griggio et al.: Moult speed affects structural feather ornaments in the blue tit (Journal of Evolutionary Biology 22:782-792, 2009).
- S. Ito och K. Wakanatsu: Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: a comparative review (Pigment Cell Research 16:523-531, 2003).
- S. J. Lin et al.: Topology of feather melanocyte progenitor niche allows complex pigment patterns to emerge (Science 340:1442-144, 2013).
- K. J. McGraw och M. C. Nogare: Distribution of unique red feather pigments in parrots (Biology Letters 1:38-43, 2005).
- R. O. Prum: Development and evolutionary origin of feathers (Journal of Experimental Zoology Mol. Dev. Evol. 285:291-306, 1999).
- R. O. Prum et al.: Two-dimensional Fourier analysis of the spongy medullary keratin of structurally coloured feather barbs (Proceedings of the Royal Society London B 266:13-22, 1999).
- D. B. Thomas et al.: Vibrational spectroscopic analyses of unique yellow feather pigments (spheniscins) in penguins (Journal of the Royal Society Interface 10:20121065, 2013).
- G. M. Toral, J. Figuerola, och J. .J. Negro: Multiple ways to become red. Pigment identification in red feathers using spectrometry (Comparative Biochemistry and Physiology Part B 150:147-152, 2008).
- M. Yu et al. The developmental biology of feather follicles (International Journal of Developmental Biology 48:181-191, 2004).

Nyckelbenet förbinder bröstbenet med skulderbladet. Det utgör hos människor och många andra ryggradsdjur skuldergördelns enda förbindelse med resten av skelettet. Hos många av oss avtecknar det sig tydligt bakom huden upptill framför bröstkorgen. Fåglarnas "önskeben" anses bestå av de två sammanväxta nyckelbenen. Nyckelbenet är ett märkligt ben dels på grund av sitt evolutionära ursprung, dels på grund av att det helt försvunnit hos många ryggradsdjur, men inte hos andra.
Bland fåglar och däggdjur tenderar nyckelbenet vara välutvecklat hos flygande och klättrande former. Det kan också vara välutvecklat hos grävande arter. Hos löpande arter är det vanligen dåligt utvecklat eller så saknas det helt. Helhetsbilden är emellertid rätt komplicerad.
Nyclelbenets bildning och funktion
De flesta av däggdjurens skelettben är ersättningsben (endokondrala ben). Ett sådant ben anläggs i form av ett brosk, format som benet, Därefter förbenas brosket. Ett fåtal skelettben är täckben (dermalben). De anläggs direkt i läderhuden, utan att ha någon broskförlaga. De är rester av det hudskelett som fungerade som ett pansar hos de många av de äldsta, nu utdöda, ryggradsdjuren. Till täckbenen hör en stor del av ansiktets skelett, inklusive underkäken, samt skalltakets ben. Hit hör, märkligt nog, också nyckelbenet (clavicula), åtminstone delvis. Det finns nämligen en del oklarheter om hur det anläggs.
Det finns två nyckelben, ett på vardera sidan av kroppen. Nyckelbenet förbinder hos däggdjur skuldergördeln med resten av skelettet. Det är kopplat till skulderbladet och bröstbenet via ledförbindelser. Detta ger ökad rörlighet åt skulderpartiet, något som kan vara en fördel för flygande, klättrande och grävande djur. Saknas nyckelbenet består skuldergördeln endast av skulderbladet och saknar förbindelse med resten av skelettet. Då underlättas förmodligen frambenens rörelser parallellt i kroppens längsriktning genom att skulderbladet blir rörligare och löpsteget därmed längre, något som är en fördel för löpande djur.
Nyckelben hos groddjur, kräldjur och fåglar
Ett nyckelben fanns hos de första, nu utdöda, fyrfota ryggradsdjuren. Hos många nutida fyrfotadjur är nyckelbenet reducerat eller så saknas det helt. Det saknas till exempel hos stjärtgroddjur (salamanderdjur) och krokodiler. Hos sköldpaddor är det inkorporerat i ryggskölden. Hos de flesta fåglar bildar de båda sammanväxta nyckelbenen det så kallade gaffelbenet ("önskebenet"; furcula). Det är emellertid osäkert om fåglarnas gaffelben verkligen härstammar från de två nyckelbenen. Det skulle kunna vara en nybildning. Alternativt härstammar det från ett annat ben i reptilernas skuldergördel. Gaffelbenet är elastiskt. Det pressas utåt när vingarna förs nedåt. Dess elasticitet anses bidra till den kraft som för vingarna uppåt. Det bidrar också till att utvidga kroppshålan när vingarna förs nedåt och därmed kanske till inandningen. Andetagen är nämligen vid flygning synkroniserade med vingarnas rörelser.

Nyckelben hos däggdjur
Inom däggdjursordningen rovdjur (Carnivora) saknar säldjuren helt nyckelben. Hos de landlevande rovdjuren är nyckelbenet reducerat eller så saknas det helt. Bland dem är det bäst utvecklat hos kattdjuren. Det ligger hos dem inbäddat i bröstmuskulaturen, men har förlorat sina ledförbindelser. Möjligen ger nyckelbenet kattdjuren en ökad rörlighet i skuldergördeln, något som skulle kunna vara en fördel när de använder framtassarna för att fånga sitt byte. Björnar saknar nyckelben. De är ju inte särskilt skickliga klättrare och härstammar förmodligen från löpande, icke klättrande djur.
De flesta gnagare (ordningen Rodentia) har nyckelben, däribland ekorren och dess släktingar bland ekorrdjuren (Sciurimorpha). Nästan alla ekorrdjur är antingen klättrare eller grävare. Tvåtåiga sengångare (släktet Choloepus) har välutvecklade nyckelben, vilket vi ju skulle förvänta oss hos klättrande djur. Men tretåiga sengångare (släktet Bradypus) saknar nyckelben. Nyckelbensanlag finns under fosterutvecklingen, men de försvinner sedan. De tvåtåiga sengångarna har dessutom välutvecklad, de tretåiga svagt utvecklad halsmuskulatur. Med tanke det likartade levnadsättet hos alla sengångare, framstår detta som synnerligen märkligt och ingen har kunnat ge en möjlig funktionell förklaring till fenomenet. Det är dock värt att notera att tvåtåiga och tretåiga sengångare inte är så nära släkt som du skulle kunna tro. De kan ha utvecklats parallellt och skaffat sig likartade anpassningar till ett liv i träden under mycket lång tid, kanske tiotals miljoner år. Till detta kommer att sengångarna, jämte manaterna, är de enda däggdjur som inte alltid har sju halskotor.
Mera förståeligt är att nyckelben saknas hos valar, som ju inte har några gångben, och hos både uddatåiga och partåiga hovdjur, som ju i regel är duktiga löpare. Likaså förståeligt är att nyckelben finns hos de flygande fladdermössen samt hos de klättrande lemurerna och aporna. Människan, som ju är en icke klättrande apa, visar inga tendenser till reduktion av nyckelbenen. Detta har kanske att göra med att det är lättare att manipulera föremål med händerna, om du har nyckelben.
Fördjupning
Hur började fåglarna flyga? Om tvåbenta dinosaurier, fjädrar och vingar Du hittar artikeln under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
Referenser
- B. K. Hall: Development of the clavicles in birds and mammals (Journal of Experimental Zoology 289:153-161, 2001).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- A. S. Romer: The vertebrate body (3rd ed, Saunders, 1962).
- S. Trotter: The significance of the "collar bone" in the Mammalia (American Naturalist 19:1172-1177, 1885).
- C. Sullivan: Floating forelimbs and clavicles (Ask a Biologist, uppdaterad 28 januari 2010).

Många djur kan ändra färg, snabbt eller långsamt. Vi skiljer på två olika slag av färgskiftning: morfologisk och fysiologisk. Morfologisk färgskiftning tar lång tid, till exempel när en hare byter från brun sommarpäls till vit vinterpäls. Fysiologiska färgskiftningar är snabba färgförändringar, som när kameleonter eller grodor byter färg.
Kromatoforer kallas de celler som orsakar fysiologiska färgskiftningar. Det finns olika slag av kromatoforer, till exempel melanoforer som är svarta eller bruna, xantoforer som är gula, erytroforer som är röda och iridoforer som reflekterar allt ljus. Många kromatoforer är stjärnformiga celler. De innehåller små pigmentkorn. Kornen kan antingen vara spridda i hela cellen eller samlade i en klump mitt i cellen. När de är spridda bidrar de till att ge djuret färg. När de är hopklumpade syns inte färgen.
Kromatoforen ändrar inte sin form vid färgbytet. I stället förflyttas pigmentkornen. Transportmekanismerna är inte helt utredda, men i vissa fall tror forskarna att pigmentkornen förflyttas längs med så kallade mikrotubuli. Mikrotubuli är små trådar som ingår i cellskelettet. Pigmentkornen transporteras längs med mikrotubuli med hjälp av så kallade motorproteiner, till exempel dynein eller kinesin. Motorproteinerna får energi till att driva rörelserna genom att spjälka den energirika molekylen ATP. Transporten av pigmentkornen styrs antingen av nerver eller av hormoner. Nervös styrning ger snabbare färgförändringar.

Bläckfiskarnas kromatoforer utgör ett undantag. I en bläckfiskkromatofor är pigmentet alltid utbrett i hela cellen. Cellen är omgiven av små muskler, placerade som uddarna i en stjärna. När musklerna är avslappade är den elastiska cellen hopdragen och så liten att dess pigment inte syns. När musklerna dras ihop, tänjer de ut cellen så att den bildar en stor rund färgad skiva. Färgen blir då synlig. Bläckfiskar kan ändra färg blixtsnabbt, tack vare att kromatoforerna utvidgas av muskelceller som i sin tur styrs av nervsystemet.
Kromatoforerna hos grodor styrs hormonellt, särskilt av ett hormon kallat MSH (melanocytstimulerande hormon) från hypofysen (undre hjärnbihanget). Forskare har studerat hur färgförändringarna styrs hos vissa grodarter. Så här kan det gå till.
Hos ljusanpassade vuxna grodor insöndras inte hormonet MSH. Då är melanoforernas svarta pigment samlat i små klumpar som knappt syns. Iridoforernas innehåll är däremot utspritt och reflekterar allt ljus som träffar huden. Allt reflekterat ljus utom det gröna absorberas emellertid av det utspridda pigmentet i xantoforerna, som ligger ovanför iridoforerna. Vi uppfattar endast det reflekterade gröna ljuset och i våra ögon blir grodan grön.
Hos mörkeranpassade grodor insöndras mycket MSH, som påverkar melanoforerna så att de svarta pigmentkornen sprids i dem. Iridoforerna dras samtidigt ihop. Dessutom skuggas iridoforerna av melanoforer. Nu reflekteras inget ljus från iridoforerna och melanoforerna absorberar allt infallande ljus, vilket leder till att grodan ser svart ut.
Unga grodyngel är svarta eftersom de insöndrar mycket MSH. Forskare har dock funnit att unga grodyngel faktiskt kan blekna under de mörka timmarna av dygnet. Detta beror på att de i mörker insöndrar hormonet melatonin från tallkottkörteln (övre hjärnbihanget). Melatoninet koncentrerar kornen i iridoforerna och i en del melanoforer.
Äldre grodyngel kan reglera sin MSH-insöndring och därigenom anpassa sin färg till bakgrunden. När de är ljusare är de svårare att upptäcka ovanför en sandbotten. De undviker därmed att bli lättfångade byten för exempelvis sländlarver. Hos vuxna grodor påverkas inga av kromatoforerna av melatonin.
Referenser
R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
P.C. Withers: Comparative Animal Physiology (Saunders College Publishing, 1992).
Förnuft, känslor och beteenden – Hur funkar hjärnan, nervsystemet, sinnen och hormoner?

Vad händer i kroppen när vi blir förälskad? Forskare har gjort flera undersökningar av den biologiska bakgrunden till förälskelse och partnerval hos människan. Vi känner också till flera biologiskt aktiva ämnen som sannolikt bidrar till känslosvallet. Detta må synas krasst. Det motsäger emellertid, som vi ska se, inte poeternas bild av kärleken. Men först lite grann om sorkar.
Monogama och promiskuösa sorkar: oxytocin, vasopressin och dopamin
Den amerikanska präriesorken (Microtus ochrogaster) är monogam. Efter den första parningen bildar hanen och honan ett par och förblir varandra trogna livet ut. Bergsorken (Microtus montanus), en nära släkting till präriesorken, är däremot promiskuös och bildar inga bestående par. Forskare har övertygande visat att skillnaden mellan de två arterna orsakas av skillnader i deras reaktioner på hormonerna oxytocin och vasopressin.
Oxytocin och vasopressin är hormoner som insöndras till blodet från bakloben i hypofysen ("nedre hjärnbihanget"). De utsöndras också från nervceller i vissa hjärndelar och fungerar där som så kallade transmittorsubstanser, genom vilka en nervcell påverkar en annan. Som hormon stimulerar oxytocin bland annat livmoderns sammandragningar vid förlosssingen och utpressningen av mjölk ut bröstvårtorna vid amning. Vasopressins hormonella effekter inkluderar ökat blodtryck och minskad urinproduktion. Men båda ämnena har också effekter på däggdjurs beteende.
Präriesorkarna har en mycket hög koncentration av en typ av receptor (mottagarprotein) för oxytocin och vasopressin på ytan av speciella nervceller i vissa delar av hjärnan. Präriesorkarnas monogama beteende utlöses av oxytocins och vasopresssins effekter på dessa hjärnregioner. Receptorkoncentrationerna i samma hjärnregioner är däremot låg hos bergssorkarna, vilket innebär att deras hjärnor är mycket mindre känsliga för oxytocin och vasopressin. Om vi blockerar receptorerna för oxytocin och vasopressin hos präriesorkarna börjar de bete sig promiskuöst. Forskarna har emellertid inte lyckats att göra bergssorkar monogama genom att injicera oxytocin och vasopressin i dem. Deras receptorkoncentrationer är så låga att hormonerna inte får någon effekt.
Hos präriesorkhanarna är det vasopressin som stimulerar till partnerval och som gör att de stannar hos sin partner. Hos honorna är det oxytocin som har dessa effekter. Båda hormonerna aktiverar hjärnans belöningssystem, som verkar via transmittorsubstansen dopamin.
Fungerar förälskade människor som sorkar?
I vilken utsträckning är sorkresultaten överförbara på människor? Det finns undersökningar som tyder på att de mycket väl skulle kunna vara det. I dessa studier har forskarna bland annat med hjälp av så kallad funktionell magnetisk resonanstomografi undersökt vilka hjärnområden som aktiveras hos förälskade par. Men det behövs mer forskning inom området och vi kan ännu inte ge något bestämt svar. Människor skiljer sig förvisso från sorkar, bland annat genom att de har högre utvecklade mentala förmågor, ett mycket avancerat socialt samspel och en högt utvecklad kultur. Allt detta gör det svårare att studera partnerval och känslomässig bindning till en partner hos människor. Nedan redovisas emellertid några intressanta resultat från studier av människor.

Forskare har vid förälskelse funnit ökad aktivitet i delar av hjärnans belöningssystem, som sannolikt innehåller höga koncentrationer av receptorer för oxytocin och vasopressin. Belöningssystemet verkar, som nämnts, via transmittorsubstansen dopamin och aktiveras i många livssituationer som ger upphov till positiva känslor. Systemet aktiveras också av alkohol, morfin, kokain och andra narkotika. Aktivering kan resultera i intensiva känslor av tillfredsställelse, lycka eller lust. Poeternas beskrivning av kärleken som ett berusande tillstånd har således en sannolik biologisk grund. Erik Axel Karlfelt (1864-1931) uttrycker det så här:
Dina ögon äro eldar och min själ är beck och kåda.
Vänd dig från mig, förr'n jag tändes som en mila innantill!
En fiol jag är med världens alla visor i sin låda,
du kan bringa den att spela, hur du vill och vad du vill.
Det finns ett stort överlapp mellan de hjärnområden som aktiveras vid förälskelse och de som aktiveras vid sex, något som inte torde vara förvånande.
Stresshormonet kortisol och transmittorn serotonin
Halterna av stresshormonet kortisol från binjurens märg är hög i de tidiga stadierna av en förälskelse, men låg i senare stadier. Detta skulle kunna vara en orsak till den stress och osäkerhet som kan märkas i början av en relation och de hälsobefrämjande effekterna av en långvarig stabil relation. Vasopressin ökar stress och rädsla, medan oxytocin minskar stress och ger en känsla av välbefinnande. Vasopressins och oxytocins effekter beror troligen bland annat på att de påverkar hypotalamus i mellanhjärnan, som styr insöndringen av kortisol, eller på att de påverkar hjärnans båda mandelkärnor (amygdala) som är av stor betydelse för upplevandet av känslor och minnen av känslor, såväl rädsla och aggression som glädje och trygghet.
Halterna av transmittorsubstansen serotonin minskar i hjärnan vid förälskelse. Detta förekommer också vid vissa psykiska sjukdomar till exempel depression, ångesttillstånd och tvångssyndrom. Depression och ångest kan för visso åtfölja en förälskelse, särskilt obesvarad sådan. Så här skriver Gustaf Fröding (1860-1911) i "Balen" efter att ha blivit avvisad av fröken Elsa Örn:
Och jag förstod, att nu var allt förbi,
och i min själ kom mörk melankoli,
jag hade dansat bort min lycka, jag,
och malört drack jag nu i långa drag,
och vart jag såg var livet dött och skumt,
och allting tölpigt, allting blygt och dumt
i trånga fjättrar åter inneslöt mig, ...
Detta är en utomordentlig beskrivning av ett depressivt tillstånd. Vid förälskelse är partnern dessutom föremål för en fixering som, mindre poetiskt uttryckt, närmar sig det tvångsmässiga. Poetiskt har detta uttryckts i "Den nya skapelsen" av Johan Henric Kellgren (1751-1795):
... Jag såg dig – och från denna dagen
jag endast dig i världen ser.
Föremålet för Kellgrens låga hette Hilma.
Förälskade människor påminner om präriesorkar
För att återvända till sorkarna, är det mycket möjligt att människor fungerar som präriesorkar. Detta skulle innebära att bandet mellan de förälskade förmedlas och stärks av oxytocin och vasopressin. Så här uttrycker Erik Lindegren (1910-1968) detta i den så ofta citerade dikten "Arioso":
Någonstans inom oss är vi alltid tillsammans,
någonstans inom oss kan vår kärlek aldrig fly
Någonstans
o någonstans
har alla tågen gått och alla klockor stannat:
någonstans inom oss är vi alltid här och nu,
är vi alltid du intill förväxling och förblandning,
är vi plötsligt undrans under och förvandling,
brytande havsvåg, roseneld och snö.
Oxytocinets mörka sida
Men oxytocin har också en annan mörkare sida. Det förstärker troligen upplevelsen av obehagliga och skrämmande sociala erfarenheter. I sådana fall skulle det till och med kunna stimulera till aggression och våld, i synnerhet mot främmande människor. Det kan vara så att oxytocin inte bara stimulerar och förstärker sammanhållningen inom en grupp, utan också främlingshat och diskriminering.
Referenser
- B.P. Acevedo, A. Aron, H.E. Fisher, och L.L. Brown: Neural correlates of long-term intense romantic love (Social Cognitive and Affective Neuroscience 7:145-159, 2012).
- A. Bartels och S. Zeki: The neural correlates of maternal and romantic love (NeuroImage 21:1155-1166, 2004).
- A. de Boer, E.M. van Buel, och G.J. Ter Horst: Love is more than just a kiss - a neurobiological perspective on love and affection (Neuroscience 201:114-124, 2012).
- M.V. Broadfoot: High on fidelity (American Scientist 90, 2002).
- C.K.W. De Dreu et al: Oxytocin promotes human ethnocentrism (Proceedings of the National Academy of Sciences 108:1262-1266, 2011).
- Y.F. Guzmán et al: Role of oxytocin receptors in modulation of fear by social memory (Psychopharmacology 231:2097-2105, 2014).
- S. Zeki: The neurobiology of love (FEBS Letters 581:2575-2579, 2007).

Påverkas människan av månen och dess faser?
Fullmånen lyser i all sin prakt. Påverkas människans fortplantning av månen och av årstiderna? Många människor har i alla tider trott att månen utövar ett inflytande på människolivet. Det handlar om allt från varulvar och mångalenskap (på engelska "lunacy", av latinets "luna", måne) till barns födelse och risken för olyckor. Det finns ett otal studier som sökt påvisa effekter av månen och dess faser på människan, men de har inte funnit några övertygande belägg för detta. Det finns dock studier som antyder att ljusa nätter med fullmåne kan påverka sömnen och leda till anfall av vissa sjukdomar, men då handlar det inte om en direkt påverkan av månen, utan om ändrade ljusförhållanden. Många djurs beteende och fortplantning styrs dock av månen. Detta gäller särskilt havsdjur, som kan ha inneboende cirkalunara rytmer, kopplade till månens faser, och cirkatidala rytmer, kopplade till tidvattnet.
Däggdjur i kalla klimat har anpassat sin fortplantning efter årstiderna. Större däggdjur har där oftast en relativt kort brunstperiod under en viss period av året, då parningen sker. Människan har levt på nordliga breddgrader i tiotusentals år. Finns det några tecken på att hon har anpassats sin fortplantning på ett liknande sätt?
Brunstperioden hos stora nordliga däggdjur är förlagd så att ungarna vanligen föds på våren. Fördelen med detta är att tillgången på föda är god under våren och sommaren. Det gör det lättare för honorna att producera mjölk och för ungarna att överleva sina första levnadmånader.
Menstruationscykler och brunstcykler
Människans menstruationscykel påminner om en brunstcykel, en regelbunden sådan med en period på cirka 28 dygn. Det finns dock skillnader. Om befruktning ej skett, kan hos många däggdjur en ny brunstcykel med ägglossning påbörjas och flera cykler kan följa efter varandra utan menstruationsblödningar. En menstruationscykel leder till en ägglossning och avslutas med en blödning, då livmoderns slemhinna till stor del stöts bort, utom då ett embryo har implanterats i den. En sådan blödning sker inte under en brunstcykel. Däremot förekommer under många däggdjurs brunstcykel en blödning från livmoderslemhinnan i samband med ägglossningen.
Har människan årstidsbunden fortplantning?
Har människan någonsin bara haft menstruationscykler med åtföljande ägglossning under en viss period av året? Detta skulle kunna ha varit funktionellt. Hos många stora däggdjur på nordliga breddgrader infaller brunsten under en period på sensommaren eller hösten, så att ungarna föds på våren, då tillgången på föda är god. Dett gäller till exempel för älgen och andra hjortdjur. Små däggdjur med kortare dräktighet fortplantar sig inte under den födofattiga vintern.

Om kvinnor hade varit mer fertila under en viss årstid, som en anpassning till ett kallt klimat, skulle födslarna på nordliga breddgrader till största delen ske under en viss period under året. Från äldre tider saknar vi data om födslarnas fördelning över året. Statistik om detta finns bara tillgängliga för de senaste århundradena, främst i länder med folkbokföring. Åtskilliga undersökningar har visat att födslarna inte är jämt fördelade över året. I stället har forskarna funnit en eller två toppar under året. Skillnaderna i födelsetal mellan årets månader varierar, men kan vara mycket stora, framför allt i u-länder. Mönstret är dock olika under olika tider och i olika delar av världen, med toppar under olika månader.
En studie av födelsetalen i Sverige, Norge och Danmark under åren 2000-2012 visade att flest barn föddes under sommaren med det största antalet i juli månad (cirka 9 procent av årets födslar) och det minsta antalet i december (mellan 7 och 8 procent av årets födslar). I dagens Sverige är således skillnaderna små. I en studie fann forskarna skillnader mellan födslarnas fördelning över året vid olika breddgrader med en topp under våren och sommaren i norr, men under hösten i söder. Skillnaden påvisades emellertid bara i USA, inte i andra länder på norra halvklotet. Det finns data som tyder på att människans fortplantning i nordliga områden påverkas av de årliga variationerna i dagslängd, möjligen på grund av årliga variationer i blodets halt av hormonet melatonin. Men endokrinologiska studier av hormoner talar emot detta. Hos många djur på nordliga breddgrader har även hanarna en brunstcykel. Testiklarna bildar då spermier bara under brunstperioden och går sedan in i ett vilstadium. Det finns ingen sådan cykel hos människan Det finns således inget stöd för tanken att vi anpassat vår fortplantning till ett liv på nordliga breddgrader.
Vad är då orsakerna till de årliga variationerna i födelsetalet? Svaret är att forskarna fortfarande inte vet. Svält under vissa delar av året är kanske en viktig förklaring i vissa u-länder och skulle kunna ha varit det i förhistorisk tid. Negativa effekter av stark värme på fortplantningen har föreslagits: hämning av ägglossningen, minskad spermieproduktion, störd implantation av embryot i livmodern och ökad dödlighet hos tidiga embryon. Men om detta vet vi inget. Födelsekontroll och en ökad konception under julferier och semester skulle naturligtvis kunna spela en roll, framför allt i i-länder. Många menar att sådana sociala och kulturella faktorer är viktigast.
Varför har människan menscykler året runt?
Människan har således sannolikt alltid haft kontinuerliga menstruationscykler året runt med en cykellängd på cirka 28 dagar. Alla hennes närmaste släktingar lever i tropikerna och har sådana menstruationscykler med åtföljande blödningar och med en ungefärlig cykellängd på 1–2 månader. Den ungefärliga längden på menstruationscykeln hos människoaporna och en annan apa anges vara:
- Schimpans: 30–35 dygn, 35–39 dygn (två studier)
- Bonobo: 42–54 dygn
- Gorilla: 26–29 dygn
- Orangutang: 27–28 dygn
- Krabbmakak: 28–32 dygn
Människans fortplantningsmönster återspeglar kanske det faktum att hon fortfarande i grunden är ett tropiskt djur. Om vi undantar områden med svåra torrperioder, är födotillgången i tropikerna ganska jämn under året. Detta gäller särskilt för människan, som under större delen av sin evolution varit allätare, jägare och samlare. Hon har dessutom kunnat öka födotillgången genom att flytta mellan olika områden. Människan har har av allt att döma inte anpassat sin fortplantning till ett liv på nordliga breddgrader med snöiga vintrar. Detta synes mig inte vara särskilt märkligt. Hon har trots allt levt i nordliga klimat i mindre är 60 000 år, en mycket kort tid ur evolutionär synpunkt. Men det hade varit mycket intressant att veta om neandertalmänniskan anpassade sin fortplantning till årstiden. Hon levde under mer än 350 000 år i Europa och överlevde flera istider. Hennes skelett visar dessutom tecken på att hon var anpassad till ett kallt klimat.
Menstruationscykler med åtföljande ägglossning kan hos människan upphöra vid svält. Detta har kanske ursprungligen varit funktionellt. Att fortplanta sig vid födobrist skulle innebära en mycket stor energiinvestering med en liten chans till ett lyckligt resultat. Både moderns och fostrets liv skulle äventyras. I vårt samhälle är det annorlunda. Om en kvinna i fruktsam ålder inte menstruerar är det ett tecken på sjukdom, såvida hon inte är gravid eller ammande. Sådan så kallad amenorré förekommer till exempel vid anorexia nervosa.

Styrs människans menscykel av månen?
En annan intressant fråga är om menstruationscykeln är kopplad till månens cykel från fullmåne till nymåne och fullmåne igen. Det är den med allra största sannolikhet inte. Människans menstruationscykel är i medeltal 28 dygn lång. Den är med största sannolikhet en inneboende frilöpande cykel, vars genomsnittliga längd råkar vara nästan en månad. Den kan påverkas av yttre faktorer, men inte av månen.
Menstruationscykelns genomsnittliga längd överrensstämmer inte med den synodiska månadens längd och den är inte synkroniserad med månens faser. Den synodiska månaden är tiden från en fullmåne till nästa, 29,5 dygn. Dessutom finns det mycket stora variationer i cykelns längd hos olika kvinnor. Olika cykler är också olika långa hos samma kvinna. Till yttermera visso har människans närmaste släktingar, schimpansen och bonobon, längre menstruationscykler, som mycket klart avviker från den synodiska månadens längd. Möjligen skulle månen kunna påverka menstruationscykeln indirekt genom sitt ljus. Nätterna är ju ljusare vid fullmåne än vid nymåne. En sådan effekt skulle i-länderna maskeras av elektrisk belysning. Forskare har emellertid studerat dogonfolket i Afrika, som inte använder elektriskt ljus. De fann inget samband mellan deras menstruationscykler och måncykeln.
Ordet menstruation är härlett från latinets menstruus som betyder månatlig. Det finns några studier som antyder att menstruationscykeln påverkas av månen, men det finns ett stort antal omfattande studier som visar på motsatsen. Det finns således inget stöd för tanken att menstruationscykeln är styrd av månen.
Referenser
- P. J. Bentley: Comparative vertebrate endocrinology (2nd ed, Cambridge University Press, 1982).
- T. K. Bauer et al.: The lunar cycle, sunspots and the frequency of births in Germany, 1920-1989 (Economics and Human Biology 11: 545-550, 2013).
- S. Bharati et al.: The effect of the lunar cycle on frequency of births, a retrospective observational study in Indian population (Indian Journal of Public Health 56:152-154, 2012).
- F. H. Bronson: Seasonal variation in human reproduction, environmental factors (The Quarterly Review of Biology 70:141-164, 1995).
- C. Cajochen et al.: Evidence that the lunar cycle influences human sleep (Current Biology 23:1485-1488, 2013)
Correspondence:
M. Cordi et al.: Lunar cycle effects on sleep and the file drawer problem (Current Biology 24:R549-R550, 2014).
M. Smith et al.: Human sleep and cortical reactivity are influenced by lunar phase (Current Biology 24:R551-R552, 2014).
C. Cajochen et al.: Reply to Cordi et al. (Current Biology 24:R795, 2014). - R. G. Foster och T. Roenneberg: Human responses to the geophysical daily, annual and lunar cycles. Review (Current Biology 18:R784–R794, 2008).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- Agnés Lacreuse et al.: Menstrual cycles continue into advanced old age in the common chimpanzee (Pan troglodytes) (Biology of Reproduction 79:407-412, 2008).
- Anders Lundquist: Cirkalunar rytm (Nationalencyklopedin, hämtad 2015-05-28).
- Anders Lundquist: Cirkatidal rytm (Nationalencyklopedin, hämtad 2015-05-28).
- M. Martinez-Bakker et al.: Human birth seasonality, latitudinal gradient and interplay with childhood disease dynamics (Proceedings of the Royal Society B Biological Sciences 281:20132438, 2014).
- J. Norum, A. Heyd och T. E. Svee: Most Scandinavians are born during summer time and less Norwegians are born the first quarter of the year, a study comparing Scandinavian birth Patterns 2000-2012 (Global Journal of Health Science 6:163-168, 2014).
- K. Shimizu et al.: Comparative study of urinary reproductive hormones in great apes (Primates 44:183-190, 2003).
- B. I. Strassman: The biology of menstruation in Homo sapiens, total lifetime menses, fecundity, and nonsynchrony in a natural-fertility population (Current Anthropology 38:123-129, 1997).

Ljudfrekvenser över 20 000 Hz definieras som ultraljud, alltså ljud som skall vara ohörbara för människor. Ju högre frekvensen är, ju högre är tonhöjden. Människans hörselområde brukar anges ligga mellan frekvenserna 20 och 20 000 Hz. Detta gäller friska unga personer. När vi blir äldre förlorar vi som bekant hörseln i det högre tonregistret. Det finns emellertid åtskilliga människor som hör ultraljud.
Unga människor kan höra ultraljud
Många personer, särskilt barn, hör högre frekvenser än 20 000 Hz. Eftersom ämnet inte är kliniskt intressant, finns det inte många undersökningar av människans hörsel vid frekvenser högre än 20 000 Hz, alltså 20 kHz. Men det finns några. I en studie fann forskarna att nästan alla försökspersonerna kunde uppfatta 22 kHz, hälften 24 kHz och en tredjedel 26 kHz. Hörseltrösklarna var dock höga, vilket innebär att det krävdes höga eller mycket höga ljudstyrkor för att personerna skulle uppfatta ljuden. Försökspersonerna var 19-25 år gamla.
Jag har fått ett e-brev från en person, som klagade över att hon kunde höra ultraljudet från den musskrämmare som hon hade installerat i sin sommarstuga. Vissa människor lär också höra ultraljudet från hundvisselpipor. Jag hittar olika uppgifter om hundvisselpipors frekvensomfång på nätet. Ofta anges 23-54 kHz. Vissa pipor anges gå ner till 18 kHz. Musskrämmare har jag inte undersökt, men de avger troligen ljud inom samma frekvensområden. Fladdermöss ekolokaliseringsläten ligger vanligen mellan cirka 20 och cirka 200 kHz. Men fladdermöss avger också läten med betydligt lägre frekvenser, som är hörbara för de flesta människor.
Det förefaller alltså inte vara alltför ovanligt att unga människor hör ultraljud inom dess lägre frekvensomfång. Själv fick jag, efter flera decenniers tystnad, höra gräshoppor igen. Men jag hade hjälp av en fladdermusdetektor, som sänkte ljudets frekvens till för mig hörbara nivåer. Jag hör inte toner högre än 8 000 Hz.
Det har hävdats att ultraljudskomponenten i musik förbättrar lyssnarupplevelsen. I en studie använde forskare balinesisk gamelanmusik. Lyssnarna kunde inte höra de toner som låg över 22 000 kHz, men de upplevde ändå musiken som mindre behaglig, när dessa toner filtrerats bort. Undersökning av deras hjärnor med positronemissionstomografi och EEG gav stöd åt slutsatsen. Men det behövs fler studier för att vi ska kunna tro på detta.
Uppfattning av högfrekventa ultraljud
Människor är emellertid känsliga för ultraljud med mycket högre frekvenser, dock bara genom direkt benledning. Det innebär att du placerar ljudkällan i direkt kontakt med skallen, vanligen pannan eller mastoidutskottet, den rundade benknölen strax bakom ytterörat. Genom benledning kan du uppfatta toner upp till cirka 100 000 Hz. Ultraljudet uppfattas som höga toner inom det hörbara området. Vi vet inte hur ultraljudet registreras. Enligt en hypotes registreras det direkt av de inre hårcellerna i början av hörselsnäckan, där de högsta tonerna inom det normala hörselområdet läses av. Enligt en annan hypotes omvandlas ultraljudet till hörbart ljud genom ett resonansfenomen inuti huvudet.
Djurskrämmare som avger ultraljud har ingen effekt
Det säljs ett flertal kommersiella apparater som uppges skrämma bort skadedjur med ultraljud. Flera studier har visat att dessa djurskrämmare inte har någon effekt på myror, kackerlackor, vägglöss, stickmyggor, råttor och möss. Tre apparater visade sig attrahera, inte skrämma bort, stickmyggor. En apparat hade till en början en svag avskräckande effekt på råttor och möss, men gnagarna vande sig snart och brydde sig inte alls om ljudet.
I några fall fungerar det med ljud som är biologiskt relevanta för en viss art. Det rör sig om varningläten, som avskräcker djuren, och parningsläten, som attraherar hanar till fällor. De flesta av dessa studier har dock gjorts i laboratoriemiljö, inte under "naturliga" förhållanden. I framtiden kan kanske apparater som avger sådana ljud bli kommersiellt tillgängliga.
Referenser
- N. Aflitto och T. DeGomez: Sonic pest repellents (Cooperative Extension, University of Arizona, 2015).
- M. Alicandri-Ciufelli et al.: Mastoid - a vestigial function in humans? (Medical Hypotheses 78:364–366, 2012).
- K. Ashihara: Hearing thresholds for pure tones above 16 kHz (JASA Express Letters 122:EL52-EL57, 2007).
- J. F. Corso: Bone-conduction thresholds for sonic and ultrasonic frequencies (Journal of the Acoustical Society of America 35: 1738-1743, 1963).
- H. G. Dieroff och H. Ertel: Some thoughts on the perception of ultrasonics by man (Archives of oto-rhino-laryngology 209:277-290, 1975).
- H. Hosoi: Activation of the auditory cortex by ultrasound (Lancet 351:496-497, 1998).
- M. L. Lenhardt: Eyes as fenestrations to the ears - a novel mechanism for high-frequency and ultrasonic hearing (International Tinnitus Journal 13:3–10, 2007).
- S. Nakagawa, C. Fujiyuki, och T. Kagomiya: Development of a bone-conducted ultrasonic hearing aid for the profoundly deaf - evaluation of sound quality using a semantic differential method (Japanese Journal of Applied Physics 52:07HF06, 2013).
- T. Nishimura et al.: Ultrasonic masker clarifies ultrasonic perception in man (Hearing Research 175:171-177, 2003).
- T. Okayasu et al.: Human ultrasonic hearing is induced by a direct ultrasonic stimulation of the cochlea (Neuroscience Letters 539:71– 76, 2013).
- T. Oohashi et al.: Inaudible high-frequency sounds affect brain activity - hypersonic effect (Journal of Neurophysiology 83:3548-3558, 2000).
- R. J. Pumphrey: Upper limit of frequency for human hearing (Nature 4222:571, 1950).

Till vänster se vi en så kallad somatosensorisk homunculus. I den återspeglar kroppsdelarnas storlek den "karta" över hudkostymen som finns i storhjärnsbarken. Ju större en kroppsdel är hos statyetten, desto större yta förfogar den över i barkens karta. Detta innebär i sin tur att statyettens stora kroppdelar har ett mycket bättre känselsinne än de små.
Till höger ser vi en motorisk homunculus. I den återspeglar kroppdelarna hur pass finstämt skelettmusklerna kan styra dem. Större kroppsdelar hos statyetten kan styras med större precision.
Vi inbillar oss gärna att vi med våra sinnen upplever en korrekt, objektiv bild av omvärlden. Men detta är mycket långt från sanningen. Vissa sinnesintryck förstärks, andra förändras och åter andra filtreras helt bort. Detta sker när vi behandlar sinnesintrycken. Hur kan vi ändå klara oss?
Sinnesintryck, stimuli, behandlas först av mottagare, receptorer, som tar emot dem, och sedan av komplicerade mekanismer i det centrala nervsystemet. Därvid förvrängs vår uppfattning av omvärlden, men den blir inte ofunktionell. Under miljontals år har evolutionen format och anpassat sinnena. Detta har lett till att vi fungerar bättre. Vår medvetna och undermedvetna upplevelse av vår omvärld är förvrängd, men detta gör att vi fungerar bättre i den miljö vi lever i. Ett roligt exempel på detta hittar vi hos grodor. I grodornas näthinna finns nervceller, som starkt reagerar på ett mycket speciellt stimulus: ett mörkt litet förmål som rör sig mot en ljus bakgrund. Dessa celler kallas "flugdetektorer". Sannolikt är grodornas världsbild starkt förvrängd till förmån för flugor. Vi människor har säkerligen många motsvarigheter till "flugdetektorer", som vi inte är medvetna om. I fortsättningen ska jag berätta om några allmänna principer för hur sinnena fungerar med känseln som exempel.
Känseln är ett viktigt sinne. Utan det skulle vi inte kunna använda våra händer med den fantastiska noggrannhet som vi är kapabla till. Utan smärta skulle vi lätt drabbas av svåra skador. Känselsinnet är emellertid inte lika väl utvecklat i alla kroppsdelar. Det är i stället evolutionärt anpassat så att vi fungerar bra. Fantastiskt nog kan det också under livets gång anpassa sig till de aktiviteter vi utför med olika kroppsdelar. Det kan med andra ord tränas.

I övrigt skiljer sig Platons filosofi i hög grad från modern naturvetenskap. Han ansåg att den verkliga världen, idévärlden, var ett ideal, överlägset de skuggor som fångarna ser. I själva verket är den värld vi upplever med våra sinnen ur vår egen synpunkt bättre än verkligheten, eftersom den underlättar vår samverkan med vår miljö. Platon ansåg också att kunskap om den verkliga världen kunde nås, inte via sinnena, utan bara genom tanken, intellektet. Detta är omöjligt. Däremot kan vi med hjälp av vetenskapliga experiment utröna hur verkligheten förvrängs av våra sinnen.
Känsel är flera sinnen
Känselsinnet är egentligen flera sinnen. Traditionellt brukar tala om känsel för smärta, tryck, lätt beröring, kyla och värme. Vi kan lägga till känsel för vibrationer. Det finns mottagare, receptorer, för alla dessa förnimmelser. Dessa receptorer är, med några undantag, specifika för en enda typ av stimulus, till exempel tryck eller smärta. De är ändar av nervfibrer, som är belägna i läderhuden, rörelseapparaten och de flesta inälvor. En del nervändar är omgivna av celler som modifierar deras svar. I rörelseapparaten och inälvorna är det glest med receptorer. Detta gör det svårt att lokalisera känselstimuli i dessa kroppsdelar. I huden finns det tätare med receptorer, vilket möjliggör en mer specifik lokalisering av stimuli.
Här kommer vi främst behandla hudens känselsinne. Men först behandlar vi några känselsinnen, som ofta inte nämns i i samband med känsel.
Kittling och klåda är särskilda sinnen
Förutom de nämnda stimulustyperna har vi kittling och klåda, som utlöses av särskilda receptorer. Dessa receptorer finns bara i huden, något som vi ska vara glada för. Klåda i inälvor skulle vi ju inte kunna lindra genom att klia. Kittling är behaglig eller obehaglig, klåda obehaglig, smärta obehaglig till olidlig. Kittlingens evolutionära ursprung är möjligen att få oss till att vifta bort blodsugande, potentiellt sjukdomsöverförande, insekter.
Proprioception, ett sinne vi inte tänker på
Slutligen finns det en typ av känselstimuli som vi inte är direkt medvetna om, från receptorer som förmedlar så kallade proprioception. Dessa mekaniska receptorer finns i muskler, senor, ledband och ledkapslar och mäter bland annat uttänjning och muskelkraft. De ger oss en uppfattning om kroppsdelarnas läge i rummet. Vi är ju medvetna om armarnas och benens läge även när vi blundar. Hjärnskadade personer, som saknar proprioception, måste titta på armarna och benen för att veta var de finns. Proprioceptorerna hjälper oss också att styra musklernas kontraktion vid kroppsrörelser.
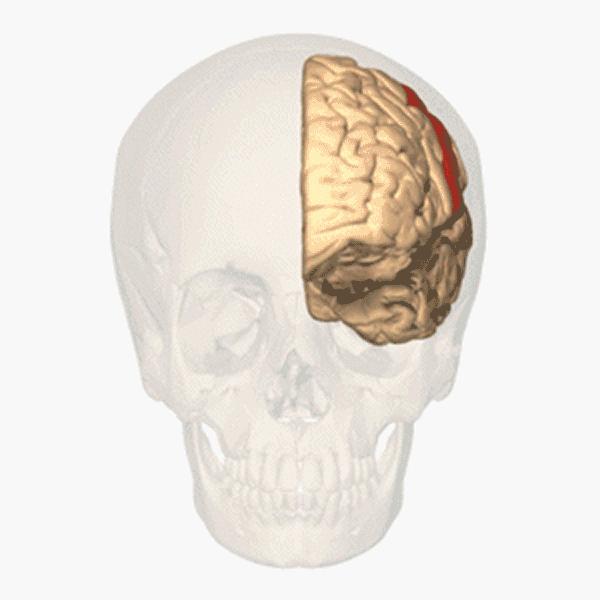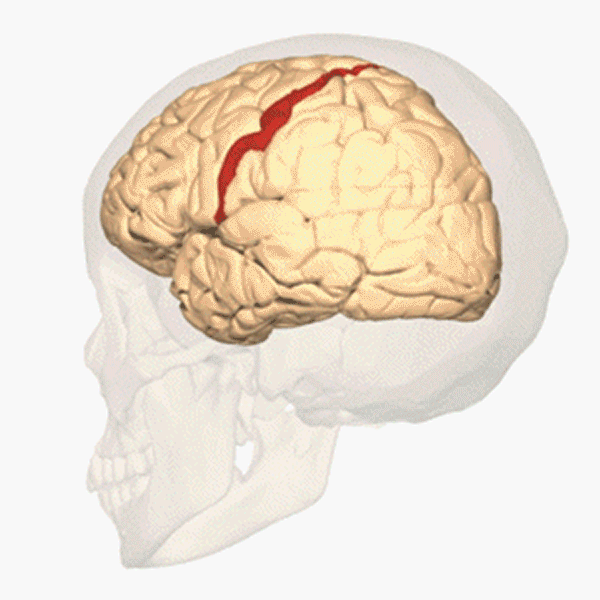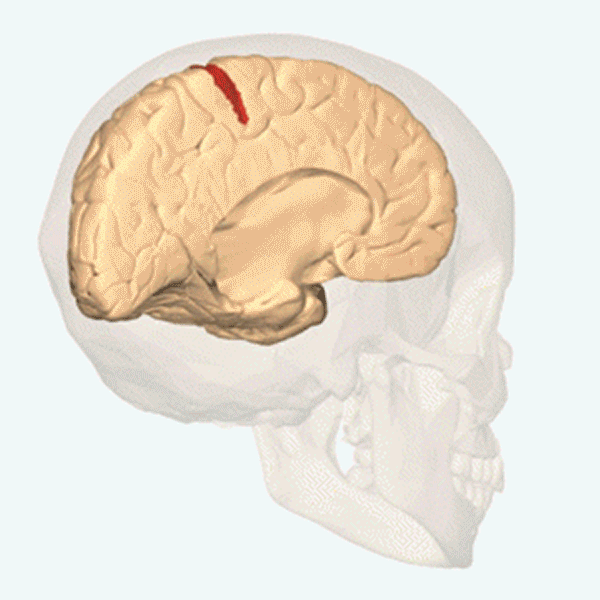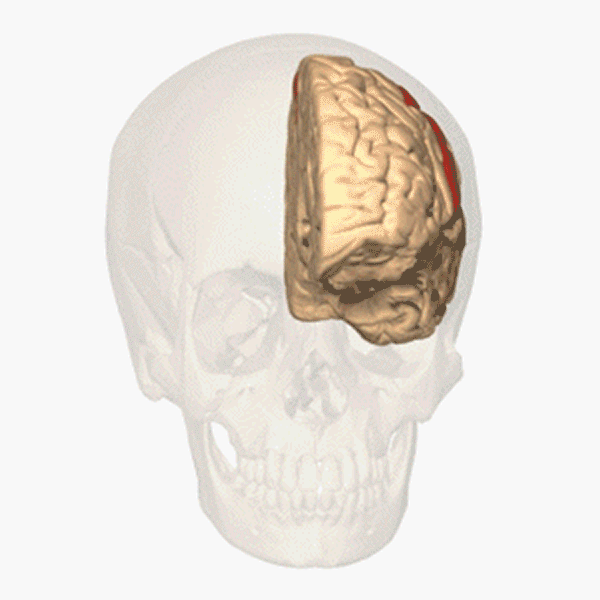
Överföring till och inom det centrala nervsystemet
Från känselreceptorerna förmedlas sinnesinformationen som nervimpulser (aktionspotentaler) först i sensoriska nervfibrer till ryggmärgen eller hjärnstammen. Där överförs informationen via så kallade synapser till andra nervceller. Dessa nervcellers utåtledande utskott (axoner) löper uppåt och bildar synapser med celler i thalamus, en del av mellanhjärnan (diencephalon). På väg till thalamus leds de över till den andra kroppshalvan. Den vänstra hjärnhalvan tar därför emot sinnesinformation från höger kroppshalva, den högra från vänster kroppshalva.
Från thalamus utgår nervfibrer som bildar synapser i storhjärnans primära känselbark. Storhjärnans bark (kortex) är dess yttersta lager med så kallad grå substans. Barken innehåller tätt packade nervcellskroppar. Innanför barken finns vit substans, som får sin färg av nervcellsutskott (axoner) omgivna av fettämnen, så kallat myelin. Där finns ocskå några kärnor med grå substans, de så kallade basala ganglierna.
Från den primära känselbarken leds information till många andra hjärndelar. Denna information bidrar, tillsammans med information från andra sinnen, bland annat till att ge underlag för att styra kroppsrörelserna samt till att ge oss en god uppfattning om vår omgivning och den rådande situationen. Den bidrar också till att aktivera relevanta känslor, till exempel smärta, skräck och välbefinnande.
Kopplingschemat avgör hur vi lokaliserar och tolkar känselintryck
En mycket viktig princip är att både sinnesreceptorer och från dem utgående nervbanor till storhjärnan måste vara specifika. Detta specifika kopplingsschema innebär att information från en receptor av en viss typ på ett visst hudområde, till exempel en smärtreceptor, alltid ska överföras till samma ställe i den primära känselbarken. Annars kan vi inte veta vilken typ av stimulus, som vi utsatts för, och vilken del av kroppen som retats. Om du elektriskt stimulerar en viss receptor, till exempel en tryckreceptor, luras hjärnan till att tolka detta som tryck på det stimulerade hudområdet. Än en gång, det är kopplingsschemat som avgör upplevelsen. Det finns dock också ospecifika nervbanor, som förmedlar information från många sinnen. De påverkar bland annat vår uppmärksamhetsgrad.
Lokaliseringen av ett känselstimulus kan dessutom skärpas genom så kallad lateral inhibition. Detta innebär förenklat att signalen från en starkt retad receptor starkt hämmar signalerna från närliggande svagt retade receptorer, medan de svagt retade receptorerna bara svagt hämmar den starkt retade receptorn. Resultatet blir att skillnaden mellan den starkt retade och de svagt retade receporernas signalstyrka blir större. Den starkt retade receptorns signal tydliggörs då.
Lateral inhibition kan förklaras med hjälp av en liknelse. Antag att det finns ett enda högt träd i en skog av obetydligt kortare träd. Antag vidare att vårt synsinne förvränger bilden och gör det höga trädet något lite lägre än det verkligen är och de låga träden väldigt mycket lägre än de verkligen är. Då skulle vi höjdskillnaden mellan träden uppfattas som större än den verkligen är och vi skulle lättare kunna urskilja det höga trädet.
Stimulus styrka kodas av nervimpulser
Styrkan på det stimulus, som en receptor utsätts för, måste översättas till nervsystemets språk. Detta innebär att ett starkare stimulus ger upphov till en högre frekvens av nervimpulser (aktionpotentialer) i den nervfiber, som receptorn ingår i, och sedan också i nervbanorna som leder till hjärnan. Ju starkare stimulus, desto fler nervimpulser per tidsenhet i nervfibrerna.
Ett annat sätt för hjärnan att mäta stimulusstyrka, förutom genom nervimpulsernas frekvens, är att summera signalerna från närliggande receptorer. Om receptorerna ligger nära varandra kommer därför fler receptorer aktiveras, så kallad rekrytering.
Allt vi upplever översätts till likadana nervimpulser
Ett filosofiskt mycket intressant faktum är följande. Alla sinnesstimuli vi utsätts för översätts till nervimpulser i nervfibrer (axoner). Hur kan vi då skilja mellan olika stimuli? För det första är det på grund av att de olika sinnesreceptorerna är specifika för och normalt bara regerar på en viss stimulustyp, till exempel ljus, ljud, doftämnen, smakämnen, mekanisk retning, kyla, värme eller smärta. För det andra är det på grund att kopplingsschemat är specifikt, vilket innebär att ljusstimuli leds till hjärnans syncentra, ljudstimuli till hjärnans hörselcentra och så vidare.
Det går dock att lura detta system om receptorer utsättes för extremt starka stimuli. Om vi får hårda slag på huvudet hör vi ljud och ser ljusfenomen. Detta påstås bero på att ögonens och öronens sinnesreceptorer aktiveras, om de utsätts för mycket starka stimuli. Det kan dock i stället vara effekter på hjärnan. Hur dessa upplevelser än uppkommer, så orsakas de av felaktiga, inadekvata, stimuli.

Känseln är evolutionärt anpassad till våra behov
De hudområden som förfogar över en stor yta i känselbarken, är försedda med mycket tätt liggande känselreceptorer. Deras stora barkyta används till att behandla den mycket utförliga information, som barken mottar från dessa hudområden. Dessa hudområden har därför en mycket väl utvecklad känsel. Detta innebär att känseln där uppvisar en mycket stor upplösning. Du kan jämföra med en datorskärm, som ju har större optisk upplösning, om pixlarna ligger mycket tätt på den.
Förmodligen behandlar större barkområden också känselinformationen mera effektivt, så att vi får en klarare uppfattning om vad det är vi känner. Dessutom är de troligen också extra känsliga för stimulus styrka, till exempel genom den ovan nämnda rekryteringen av receptorer.
I människans sensoriska homunculus är framför allt huvudet och händerna gigantiska. Dessa kroppsdelar förfogar således över de största ytorna i den primära känselbarken och har därmed det bästa känselsinnet. Ett välutvecklar känselsinne för huvudet, särskilt läpparna och tungan, finns sannolikt hos alla däggdjur. Detta underlättar födointag och undersökning av omvärlden med nos och morrhår. Dessutom är däggdjurens ansikte, till skillnad från andra ryggradsdjurs, försett med så kallad mimisk muskulatur. Sådan muskulatur möjliggör kommunikation med ansiktsuttryck. Dessutom förbättrar de flesta däggdjur, dock ej människan, sin hörsel genom att med sina ansiktsmuskler vrida öronen mot ljudkällor.
Den extremt välutvecklade känseln i händerna är däremot troligen unik för människan. Under vår evolution har händerna utvecklats till fantastiskt känsliga redskap. Denna egenskap är kanske lika viktig för oss som vår välutvecklade kognitiva förmåga ("intelligens").
Hjärnan anpassas till behovet under livets gång
Förr trodde forskarna att de sensoriska barkytorna för de olika kroppdelarna var fastlagda från födseln, Numera vet vi att inte bara dessa områden, utan hela storhjärnan, kan anpassa sig till våra behov under loppet av vårt liv, särskilt hos barn och ungdomar, men även hos vuxna. Hjärnan kan alltså tränas, precis som rörelseapparaten. Hjärnan plasticitet är mycket större än vi kunde ana. Händernas barkområden är till exempel mycket större än normalt hos pianister som börjat träna i unga år. Detsamma gällde kanske de avlägsna förfäder till oss som lärde sig göra fantastiskt användbara stenredskap. Det mest extrema exemplet på hjärnans plasticitet är att läkare hos barn med mycket svår epilepsi kan operera bort hela hjärnbarken i den ena storhjärnshalvan med förvånande små effekter.
Referenser
- K. E. Barnett, S. M. Barman, S. Boitano, och H. L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (4th ed, Sinauer, 2018 ).
- Platon: Staten (Atlantis, 2017; översatt av Jan Stolpe).
- G. J. Tortora och M. T. Nielsen: Principles of human anatomy (12th ed, Wiley, 2012).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).

Andra hypoteser är att ränderna gör insekter mindre bitbenägna, att ränderna gör att zebrorna lättare känner igen varandra och att ränderna underlättar kylning av zebrorna päls. Såvitt forskarna nu vet, är skydd mot bitande insekter den sannolikaste hypotesen. Men det behövs fler studier och vi kan inte utesluta att ränderna har flera funktioner.
Varför har zebror ränder? Det är en fråga som forskare har ställt sig i mer än hundra år, utan att kunna besvara den. Först under 2000-talet har de på allvar angripit den i en rad vetenskapliga studier och börjat närma sig svaret.
Frågan kan besvaras på två sätt beroende på vad du menar med ordet varför.
Vilken är den evolutionära fördelen med ränder?
För det första kan du med ordet varför fråga sig vilken funktion ränderna har, det vill säga vad som har gynnat randiga djur under hästsläktets evolution i Afrika. Det finns flera hypoteser.
En hypotes är att ränder gör det svårare för rovdjur, i zebrornas fall framför allt lejon och hyenor, att se bytesdjuren. Zebrorna kanske smälter ihop med bakgrunden så att lejonen har svårt att se dem. Detta må synas egendomligt för oss människor som ser zebrorna så tydligt, men du får tänka på att lejon och hyenor inte ser världen på samma sätt som vi. Bland annat så har de inte vårt välutvecklade färgseende. Alternativt så försvårar randningen lejonens avståndsbedömning eller så gör randningen det svårare för lejonen att urskilja en enskild zebra bland alla de andra zebrorna.
Enligt en annan hypotes gör randningen bromsar och tsetseflugor, kanske också andra blodsugande insekter, mindre benägna att sticka zebrorna. Denna hypotes stöds av försök. Modellförsök har visat att bromsar är mindre benägna att bita randiga ytor. Tsetseflugor föredrar stora, helt mörka djur. Bromsar sprider flera sjukdomar. Tsetseflugor sprider sjukdomar som kallas trypanosomoser till zebror och andra djur, inklusive människor. Stickande insekter orsakar dessutom blodförluster och andra hälsoproblem.
Enligt en tredje hypotes har ränderna en social funktion. Eftersom randningen är olika hos olika individer, så skulle ränderna till exempel kunna fungera som igenkänningstecken inom flocken.
Enligt en fjärde hypotes skulle de olikfärgade ränderna vid uppvärmning av pälsen ge upphov till luftvirvlar som underlättar kylning av huden i en varm miljö.
Sammanfattningsvis kan vi säga att ingen av de ovan redovisade hypoteserna ännu kan uteslutas. Det behövs fler fältstudier av zebror, av deras predatorer och av stickande insekter i djurens naturliga miljö. Hypotesen att ränderna lurar stickande insekter har dock starkast stöd. Men de olika hypoteserna utesluter inte varandra. Zebrornas ränder kan alltså ha flera funktioner.

Hur uppkommer ränderna under fosterutvecklingen?
För det andra kan du med ordet varför fråga dig hur randningen uppkommer under zebrornas fosterutveckling, det vill säga vilka som är de bakomliggande orsakerna till randigheten. Hudens pigmentceller härstammar från den så kallade neurallisten som anläggs under embryonalutvecklingen. Nervsystemet anläggs som en inbuktning av huden längs med embryots mittlinje på ryggsidan. Inbuktningen sluts sedan till ett rör, nervröret. I samband med nervrörets bildning uppkommer de båda längsgående neurallisterna på båda sidor om nervröret under huden. Neurallisternas celler vandrar ut i kroppen och ger upphov till en rad strukturer, bland annat större delen av det perifera nervsystemet och, som sagt, hudens pigmentceller. Pigmentcellernas pigmentkorn byggs sedan in i hudens hornlager och i pälsens hår. På så sätt får hud och päls sina färger. Hos zebrorna kommer pigmentcellerna påverkas så att embryots päls indelas i omväxlande pigmenterade svarta och pigmentfria vita zoner. Pigmentcellerna i de svarta ränderna bildar svart pigment, som byggs in i hårstråna. I de vita ränderna hämmas pigmentcellernas pigmentbildning, vilket innebär att hårstråna helt saknar pigment. Forskarna vet mycket lite om hur detta går till.
Är zebrorna vitrandiga eller svartrandiga?
Är zebrorna vitrandiga eller svartrandiga? Eftersom de vita och svarta ränderna ofta är ungefär lika breda, kan vi med fog hävda att denna fråga språkligt är meningslös. Zebrorna är helt enkelt svart-och-vit-randiga. Men det finns faktiskt argument som talar för att de är svartrandiga och argument som talar för att de är vitrandiga.
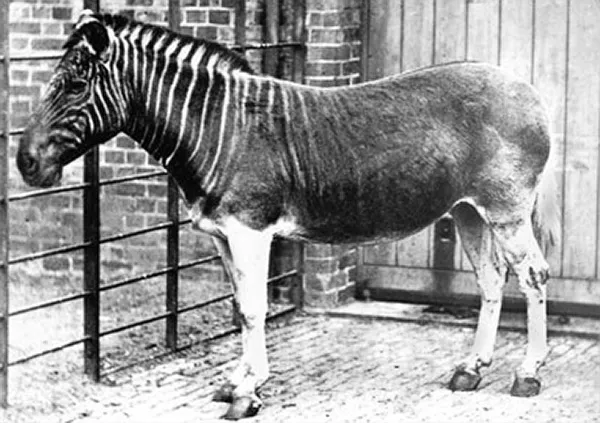
Om du jämför med andra hästdjur, skulle du kunna dra slusatsen att zebrorna är mörkrandiga mot ljus botten. Zebrornas nu levande släktingar bland hästdjuren är Przewalskis häst (Equus przewalskii), den afrikanska vildåsnan (Equus africanus), kiangen (Equus kiang) och den asiatiska vildåsnan (Equus hemionus). Dessa arter har en ljus färg på rygg och flanker: grå, gråbrun, brun eller rödbrun. Stora delar av kroppen är oftast vita, framför allt buken. Ett undantag är kiangen, vars vinterpäls vanligen är mörkbrun på rygg och flanker, dock långt ifrån svart. Den ena underarten av den afrikanska vildåsnan har tunna zebraliknande svarta streck mot ljus botten på benen. I övrigt är ingen av arterna randiga. Flera av arterna har dock ett mörkt streck längs med ryggen, en så kallad ål. Ser vi tiill zebrorna, kan vi notera att två av de tre zebraarterna, grevyzebran (Equus grevyi) och bergszebran (Equus zebra), har markant vit buk. Hos den vanliga zebran, (Equus burchelli), tendera dock ränderna att mötas på buken Benens insidor kan dock vara mer eller mindre vita hos denna art och varianter med vit bakkropp förkommer ibland. Sammantaget, kan allt detta tolkas som om den ursprungliga färgen hos hästdjuren var ljus med helt vita inslag och att svarta ränder har upkommit hos zebrorna.

Men det har också hävdats att zebrorna tvärtom är vitrandiga mot mörk botten. Ett argument för detta skulle vara de tunna svarta eller grå ränder som hos den vanliga zebran uppkommer i mitten av vita ränder, om de senare är mycket breda. Detta har tolkats så att någon slags hämning av pigmentbildningen bli för svag om de vita ränderna är för breda och att grundfärgen skulle vara mörk. Pälsmönstret hos den utdöda kvaggan har tolkats som om den var ljust randig mot mörk botten.
Huden under zebrornas päls svart. Även detta kan tolkas så att djuren är vitrandiga mot en mörk botten. Men många däggdjur och fåglar har en svart hud under en ljus päls eller fjäderdräkt. Ett exempel är isbjörnen. Ingen skulle få för sig att kalla den svart. Det är emellertid mycket märkligt att ett tropiskt djur, som zebran, har svart hudfärg. Detta borde öka dess värmeupptag genom att öka absorptionen av det synliga ljus som passerar genom den tunna pälsen.

Den utdöda kvaggan: mörkrandig eller ljusrandig?
Kvaggan (Equus quagga quagga), en utdöd sydafrikansk zebraunderart, uppvisade stora variationer i färg och randning, se bilderna ovan. Den var oftast randig huvudsakligen på huvudet och framkroppen. Men ibland var den randig över hela kroppen. De ljusa ränderna var ofta tunna. Forskarna anser vanligen att den framtill var ljusrandig mot en brun bottenfärg, påminnande om den enfärgade bakkroppens färg. Detta framgår av de få svartvita fotografier som bevarats. Å andra sidan ger flera samtida illustrationer och några av de bevarade uppstoppade djuren det motsatta intrycket. När det gäller de uppstoppade djuren skulle detta kunna bero på blekning av färgerna.
The Quagga Project (extern webbplats) försöker nu "återskapa" kvaggan genom att korsa lämpliga zebror tillhörande en närbesläktad underart. De försöker få fram individer med samma pälsmönster som kvaggan hade. Men detta innebär inte att de återupplivar kvaggan, bara att de avlar fram zebror med ett likartat pälsmönster.
Fördelar med att vara randig – utförlig diskussion
I en studie från 2012 visade forskare genom experiment att zebraliknande randning gör en yta mindre attraktiv för bromsar. Bromsar är blodsugande flugor inom familjen Tabanidae som angriper både hästar och zebror. De attraheras mer till svarta än till vita hästar. I försök med svarta, vita och randiga hästmodeller fann forskarna att bromsarna var allra minst attraherade till randiga modeller, vars ränder var av ungefär samma bredd som zebrornas. Forskarna fann också att det reflekterade ljusets polariseringsegenskaper hos hästmodellerna överensstämde väl med egenskaperna hos zebraskinn. Det reflekterade ljusets polarisering är av stor betydelse för bromsarnas beteende, när de söker efter djur att suga blod från. I två senare undersökningar har dessa resultat bekräftats.
Forskarna skulle gärna vilja att fler försök gjordes med zebror i deras naturliga afrikanska miljö samt att försöken omfattade både bromsar och tsetseflugor. Icke desto mindre, stödjer dessa studier hypotesen att zebrornas randning skyddar dem från blodsugande insekter.
I en annan studie har forskarna genom datasimuleringar visat att randningen skulle kunna ge upphov till synvillor hos de rovdjur som ser zebrorna röra sig. Sådana optiska illusioner kan ge betraktaren uppfattningen att zebrorna rör sig i en annan riktning än de verkligen gör, så kallat rörelsekamouflage. Synvillorna kan tänkas lura såväl lejon och andra rovdjur som tsetseflugor, bromsar och andra insekter. Men detta har inte bekräftats genom direkta observationer i naturen.
I en studie från 2016 beräknade forskare mått på förmågan att med synen urskilja kontraster hos zebror samt deras viktigaste predatorer, lejon och fläckig hyena. De beräknade dessa mått för tre olika ljusstyrkor, motsvarande dag, skymning eller månljus natt samt månfri natt. Forskarna utgick emellertid från data från tamkatt. De drog slutsatsen att zebrorna inte vara svårare att urskilja för predatorerna än andra bytesdjur. Men inte heller detta har bekräftats genom direkta studier.
I en omfattande studie från 2014 mätte forskare graden av randighet med flera olika mått hos vilda hästdjur (zebror, halvåsnor, åsnor och Przewalskis häst). De tog också fram mått på risken att bitas av bromsar och tsetseflugor i olika delar av världen. Forskarna fann hästdjuren var "randigare" i de områden som var svårast plågade av bromsar och tsetseflugor. De fann inget stöd för hypoteserna att randighet skulle göra det lättare att undkomma rovdjur, att den skulle underlätta kylning av huden eller att den skulle ha en social funktion. Detta stärker ytterligare hypotesen att zebrornas randning skyddar dem från blodsugande insekter, men du måste ha i minnet att studien visar på korrelationer, om än starka sådana, inte orsakssamband.
I en studie från 2016 fann forskare visst stöd för tanken att andra egenskaper hos pälsen skulle skydda mot bitande insekter, till exempel de ovan nämnda synvillor som ränderna kan ge upphov till. Studien var emellertid teoretisk och byggde på ögats egenskaper hos spyflugor, inte hos stickande insekter.
I en studie från 2015 fann forskare ett samband mellan rändernas mönster och omgivningens temperatur. De utgick från fotografier av en zebraart i olika delar av dess utbredningsområde. Forskarna drog slutsatsen att ränderna underlättar avkylning av huden. De svarta ränderna värms till högre temperaturer än de vita av det synliga solljuset. Luften närmast ovanför de svarta ränderna kommer därför att bli varmare än luften ovanför de vita. Eftersom varm luft stiger snabbare uppåt, ju varmare den är, kommer luften att röra sig snabbare över de svarta än över de vita ränderna. Enligt temperaturhypotesen skulle detta kunna ge upphov virvlar av kylande luft närmast pälsen. Denna hypotes testades experimentellt i en studie publicerad 2018. Forskarna täckte vattenfyllda tunnor med päls från zebra, nötboskap och häst. De mätte temperaturen mitt inne i tunnorna medan de förvarades utomhus under fyra månader. Den inre temperaturen svarar mot temperaturen i kroppens kärna. Eftersom däggdjur reglerar kärntemperaturen, inte hudens temperatur, är den ett bättre mått än hudtemperaturen. Dessa försök stödde inte kylningshypotesen. Tunnorna med zebraskinn blev inte svalare än tunnor med grått hästskinn. Det finns också andra invändningar mot kylningshypotesen.
Om det är en stor fördel med att vara randig, kan vi fråga oss varför inga andra av savannens djur är så markant randiga som zebrorna. En förklaring, som framkastats, är att det beror på att zebrorna har tunnare päls än många andra savanndjur. Därmed skulle de vara mindre skyddade från stickande insekter och mindre isolerade från den infallande solstrålningen. Å andra sidan finns det många savanndjur med två ganska starkt kontrasterande pälsfärger, till exempel leopard, gepard, hyenor och flera antiloparter. Dessa djur är antingen randiga eller fläckiga. Girafferna uppvisar en mycket markant färgkontrast mellan fläckarna och angränsande områden. Det är dock oklart om dessa pälsmönster utgör kamouflage eller om de har någon annan funktion.
Skydd mot bitande insekter framstår nu som den sannolikaste fördelen med zebrornas ränder, men det sista ordet är inte sagt i frågan. Det behövs fler studier av levande zebror i deras naturliga miljö, men sådana är inte alltid lätta att utföra. Och de olika hypoteserna utesluter inte varandra.
Fördjupning
Fåglarnas färgrika fjädrar och däggdjurens hårstrån: hur de bildas och får sin färg. Pigment och strukturfärg. Artikeln finns under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?
Referenser
- M. Blahó et al.: Stripes disrupt odour attractiveness to biting horseflies: battle between ammonia, CO2, and colour pattern for dominance in the sensory systems of host-seeking tabanids (Physiology & Behavior 119:168-174, 2013).
- K. H. Britten, T. D. Thatcher, och T. Caro: Zebras and biting flies: quantitative analysis of reflected light from zebra coats in their natural habitat (PLoS ONE 11:e0154504, 2016).
- T. Caro et al.: The function of zebra stripes (Nature Communications 5:3535, 2014).
- Á. Egri et al.: Polarotactic tabanids find striped patterns with brightness and/or polarization modulation least attractive: an advantage of zebra stripes (Journal of Experimental Biology 215:736-745, 2012).
- Encyclopedia of Life (hämtad 6 april 2017).
- S. F. Gilbert: DevBio.com (hämtad 10 april 2017).
- M. Gosling: Biting flies, lions, and the evolution of zebra stripes (Ecology 98:2227-2229, 2017).
- M.D. Hofmeyr: Thermal properties of the pelages of selected African ungulates (South African Journal of Zoology 20:179-189, 1985).
- Gábor Horváth et al.: Experimental evidence that stripes do not cool zebras (Scientific Reports 8:9351, 2018).
- M. J. How och J. M. Zanker: Motion camouflage induced by zebra stripes (Zoology 117:163-170, 2014).
- B. Larison et al.: How the zebra got its stripes: a problem with too many solutions (Royal Society Open Science 2.1:140452, 2015).
- R. Lydekker: Note on the skull and markings of the quagga (Proceedings of the Quarterly Journal of the Geological Society 26:426-431, 1904).
- A. D. Melin et al.: Zebra stripes through the eyes of their predators, zebras, and humans (PLoS ONE 11:e0145679, 2016).
- G. Vaduva: The evolution of striped coat patterns in African mammals - what function do the stripes have? (Journal of Biology and Life Science 7:53-70, 2016).
- J. K. Waage: How the zebra got its stripes - biting flies as selective agents in the evolution of zebra coloration (Journal of the Entomological Society of South Africa 44:351-358, 1981).

Det finns många ryggradslösa djur med mer än två ögon. Men alla ryggradsdjur har väl två ögon? Nej, det finns faktiskt några ryggradsdjur med ett tredje öga. Ett av dem är tuataran, djuret på bilden ovan. Den är berömd för att ha ett tredje öga mitt på huvudets ovansida, ett så kallat hjässöga.
Forskare har spekulerat att de allra tidigaste ryggradsdjuren hade fyra ögon. Förutom det ögonpar som alla ryggradsdjur har i dag, så hade de kanske ett par uppåtriktade ögon på huvudets ovansida, möjligen ett till vänster och ett till höger. Med dessa hjässögon kunde dessa urryggradsdjur kanske upptäcka fiender i vattnet ovanför dem eller styra sina kropprörelser.
Många av de tidigaste vattenlevande ryggradsdjuren under devonperioden (för cirka 415–360 miljoner år sedan) hade faktiskt ett tredje öga mitt på hjässan. Sådana hjässögon fanns både hos de så kallade pansarrundmunnarna och hos många tidiga benfiskar. Hos de fyrfota landlevande ryggradsdjurens förfäder, de köttfeniga fiskarna, var de vanliga. Ett sådant tredje öga fanns kvar hos många av de tidiga groddjuren och reptilerna, bland annat hos therapsiderna (en grupp "däggdjursliknande reptiler"), våra egna förfäder. Men sedan försvann hjässögonen hos de flesta ryggradsdjur. Hjässögon finns i dag kvar hos nejonögonen, hos några fiskar, hos en del groddjur, hos några ödlor och hos tuataran. Tuatarans hjässöga är tillbakabildat, men det är försett med såväl hornhinna och lins som näthinna. Troligen producerar det ingen bild, utan registrerar bara ljusintensiteten. Detsamma gäller förmodligen för de andra nutida hjässögonen.

Hjässögon styr dygnsrytmen
Hjässögonen är emellertid känsliga för ljus och mörker. Vi vet inte mycket om deras funktioner. Men de mäter troligen dagslängden och korrigerar därmed djurens biologiska klockor. Alla djur har biologiska klockor med en inneboende dygnsrytm (cirkadiansk rytm), som är oberoende av yttre stimulering. Men klockorna tenderar att gå fel och då kan de påverkas av yttre stimulering, oftast dagsljus, som ställer klockorna rätt. Ljuset fungerar då som tidgivare.
Men varför tror forskare att det funnits inte ett, utan två hjässögon? Både hjässögonen och de vanliga ögonen bildas under embryots utveckling som utbuktningar från mellanhjärnan (diencephalon), den andra av hjärnans fem huvuddelar framifrån räknat. Hjässögonen bildas från mellanhjärnans tak. Det märkliga är att det hos ryggradsdjuren förekommer flera olika utbuktningar i mittlinjen från mellanhjärnans tak. Två sådana utbuktningar är pinealorganet och parapinealorganet. Hos nejonögonen bildar både pinealorganet och parapinealorganet ögonliknande strukturer, men pinealögat är bäst utvecklat. Hos groddjur bildas hjässögat troligen av pinealorganet och benämns frontalorgan. Hos tuataran och de hjässögonförsedda ödlorna utgörs hjässögat av parapinealorganet och kallas parietalöga.
Ljuskänsliga parapinealorgan och pinealorgan
De flesta ryggradsdjur som saknar hjässögon, har ett ljuskänsligt parapinealorgan eller pinealorgan under skalltaket. Många av dessa organ har förmodligen samma funktion som hjässögonen, alltså att med hjälp av ljus som tidgivare ställa om en överordnad biologisk klocka i hjärnan. Det handlar då om ljus som passerar igenom skalltaket. Hos en del av organen har forskare också visat att de insöndrar hormonet melatonin, som bidrar till att styra dygnsrytmen. I andra fall har organen kanske förlorat sin funktion. Hos vissa ryggradsdjur har forskarna inte hittat några sådana organ.
Hos fåglar registreras ljus som passerar genom skalltaket av celler av pinealorganet. Hos höns har forskare visat att dessa celler känner av dygnets växlingar mellan ljus och mörker. Ljuset fungerar som tidgivare för biologiska klockor. En blind höna kan inte hitta ett korn, men den kan fortfarande korrigera sin dygnsrytm.

Däggdjurens tallkottkörtel och hormonet melatonin
Pinealorganet har hos de däggdjuren, inklusive människan, omvandlats till en körtel som ligger innanför skalltaket. Den kallas tallkottkörteln eller epifysen och producerar bland annat hormonet melatonin. Däggdjurens tallkottkörtel innehåller inga ljuskänsliga celler. Hos däggdjuren är det i stället vissa nervceller i ögat som reagerar på det tidgivande ljuset med hjälp av det ljuskänsliga pigmentet melanopsin. Notera att dessa celler är nervceller. Syncellerna, det vill säga stavarna och tapparna, påverkar inte våra biologiska klockor. Hos däggdjur förmedlas de tidgivande dygnsrytmssignalerna till en parig kärna i hypofysen, en del av mellanhjärnan. Kärnan kallas nucleus suprachiasmaticus och fungerar som en överordnad biologisk klocka, som påverkar andra organ i kroppen. Den styr bland annat insöndringen av hormonet melatonin från tallkottkörteln. Värt att notera är att det finns många biologiska klockor i kroppen, kanske i varje organ eller till och med i varenda cell. Dessa klockor styrs till stor del av den överordnade klockan i nucleus suprachiasmaticus. Men många av dem kan sannolikt också ticka självständigt, styrda av andra tidgivare än ljuset.
Hormonet melatonin har påvisats hos bland annat groddjur, fåglar och däggdjur. Hos grodor kan detta hormon åstadkomma färgväxling genom att påverka vissa celler i huden. Hos människor är melatonin ett natthormon som insöndras under den mörka tiden av dygnet. Som nämnts styrs insöndringen av melatonin av den överordnade klockan i nucleus suprachiasmaticus. Melatoninet i sin tur styr en rad dygnsrytmer i kroppen, till exempel växlingen mellan vakenhet och sömn. Det har också föreslagits att puberteten utlöses av minskande melatoninhalter hos barnet, men det finns fakta som talar emot detta.
Den store 1600-talsfilosofen Descartes ansåg att själen samverkar med kroppen via tallkottskörteln. Han ansåg också att själen inte är materiell, utan att den existerar skild från kroppen och opåverkad av naturens lagar. Denna teori är förstås numera förkastad. Men mycket är ännu oklart när det gäller tallkottskörtelns och melatoninets roll hos människan och andra djur. Descartes gjorde bestående insatser inom matematik och optik, notera ljusets brytning i ögonlinserna på bilden ovan.
Referenser
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).
- P. Ekström och H. Meissl: Evolution of photosensory pineal organs in new light, the fate of neuroendocrine photoreceptors (Philosophical Transactions of the Royal Society London B 358:1679-1700, 2003).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. Okano et al.: Diversity of opsin immunoreactivities in the extraretinal tissues of four anuran amphibians (Journal of Experimental Zoology 286:136-142, 2000).
- I. R. Schwab och G. R. O'Connor: The lonely eye (British Journal of Ophthalmology 89:256, 2005).
- S. Tamotsu och Y. Morita: Photoreception in pineal organs of larval and adult lampreys, Lampetra japonica (Journal of Comparative Physiology A 159:1-5, 1986).
- Béla Vigh, I. Vigh-Teichmann, K. Debreceni och J. Takács: Similar fine structural localization of immunoreactive glutamate in the frog pineal complex and retina (Archive of Histology and Cytology 58:37-44, 1995).

Däggdjurens päls är nästan aldrig bjärt färgad. Med få undantag är den brun eller grå. Ibland är den svart eller vit. Fåglar har däremot ofta en bjärt färgad och brokig fjäderdräkt. Sannolikt beror detta på att de flesta däggdjur är nattaktiva med ett dåligt färgseende. Dessutom är det mycket få djur som kan se färger i svag belysning. De flesta fåglar är däremot dagaktiva med ett välutvecklat färgseende. Men varför är fåglar, som ser färger så bra, så ofta vita?
Fjäderdräkt som kamouflage
Färgad fjäderdräkt hos fåglar fungerar ofta som kamouflage (krypsis). Hos många arter är fjäderdräkten färgad i bruna och grå nyanser, som ger ett mycket gott kamouflage. Ofta är det bara honorna, som är kamouflerade, något som kommer väl till pass när de ruvar äggen. Buken är ofta ljusare än ryggen. Detta gör djuren svårare att upptäcka.

Varför har många hanar färgstark fjäderdräkt?
Det finns emellertid också många fåglar som är bjärt färgade. Röda, gula, gröna och blå färger är vanligt förekommande. Färgerna kan ha en funktion vid kommunikation med artfränder. Ofta är det bara hanarna som bär på iögonenfallande färger. Då handlar det om så kallad sexuell selektion. Hanar med klara färger signalerar att de är starka och fortplantningsdugliga. Sådana hanar kan skrämma bort rivaler. De kan också vara särskilt attraktiva för honorna. Välkända exempel på sexuell selektion är fasan och tjäder. Paradexemplen är paradisfåglarna på Nya Guinea och påfågeln.
Kan vit fjäderdräkt ge kamouflage?
Redan Charles Darwin frågade sig, varför så många fåglar är vita. Vita fjädrar saknar färgämnen. De innehåller en mängd mycket små luftbubblor som reflekterar ljuset i alla riktningar, så kallad spridning. Alla färger inom det för människan synliga spektralområdet reflekteras i lika hög grad. Detta gör att fjädrarna ter sig vita för våra ögon. Vitt ljus är ju en kombination av alla regnbågens färger. Fjädrarnas struktur bidrar också till deras vita färg. Det handlar om samma fenomen som gör mjölk vit, fast i mjölken sprids ljuset av små fettdroppar.
Hos vita fåglar har båda könen ofta likfärgad fjäderdräkt. Därmed är sexuell selektion mindre sannolik. I kalla snötäckta områden ger den vita fjäderdräkten kamouflage. Detta gäller förmodligen ripor, fjälluggla och möjligen de svanarter, som häckar i Arktis. Men det finns många vita fåglar i andra klimatområden, även i tropikerna. Exempel är flera svanarter, albatrosser, de flesta måsfåglar och flera hägerarter.
Det är anmärkningsvärt många havsfåglar som är vita, åtminstone på undersidan. Det har föreslagits att de därigenom bli svårare att upptäcka för de fiskar som är deras byten. Men det finns tunga invändningar mot denna hypotes. De kamoufleras möjligen mot en bakgrund av vita moln, men inte mot en gråmulen eller blå himmel. Fåglar som simmar på vattnet ses av fiskar, oberoende av vilken färg de har på undersidan.

Kan vit fjäderdräkt gynna temperaturreglering?
Ljus och mörk fjäderdräkt kan emellertid ha betydelse för en fågels upptag av värme genom absorption av synligt ljus från solen. Men detta gäller bara vid direkt solbelysning. I molnigt väder och i skugga står det synliga ljuset för en mycket liten del av värmeupptaget.
Den för människan synliga solstrålningen kan reflekteras av, absorberas av och/eller passera igenom en fågels fjäderdräkt. Forskare måste emellertid undersöka arten i fråga för att ta reda på vad som händer med ljuset. Det är viktigt att framhålla att det inte bara är fjädrarnas färg som avgör hur ljuset beter sig. Även fjädrarnas struktur och fjäderdräktens täthet är av betydelse. Notera att jag här först bara talar om för människan synlig solstrålning, inte om infraröd strålning ("värmestrålning") och UV-strålning. Resonemangen gäller också däggdjur med vit päls.
En vit fjäderdräkt hos en fågelart kan tänkas reflektera merparten av den synliga solstrålningen och absorbera resten nära ytan, varvid ytan värms upp. Om vinden blåser eller om djuret rör sig, kommer en stor del av den på ytan bildade värmen att övergå till luften genom så kallad konvektion. Då blir fågelns värmeupptag från solen litet, och risken för överhettning minskar i varmt väder.
En vit fjäderdräkt hos en annan art kan tänkas släppa igenom en stor del av den synliga solstrålningen ända ner till huden och reflektera en mindre del på fjäderdräktens yta. Om huden är svart, absorberar den nästan all synlig strålning som når den och reflekterar mycket lite synlig strålning. Då blir värmeupptaget stort från den synliga strålningen och relativt lite värme försvinner från djuret genom konvektion under blåsiga förhållanden. Risken för nedkylning i kallt blåsigt väder minskar då. Intressant nog finns det flera vita fåglar som har svart hud under den vita fjäderdräkten.
Vi kan mycket grovt säga att solstrålningsenergin under en klar himmel vid jordytan till 40 procent består av för människan synligt ljus och till 60 procent av infraröd strålning. Ur uppvärmingssynpunkt kan du försumma andra våglängder, inklusive UV-strålning. Men UV-strålningen har naturligtvis andra effekter. Den är till exempel kancerframkallande. Hos människor stimulerar den också en kemisk reaktion i huden, som gör att vi själva kan bilda vitamin D.

De flesta fåglar kan, intressant nog, se UV-strålning. För dem ingår alltså UV-strålningen i det synliga ljuset. Hos vissa vita fåglar, till exempel gråtrut och fjälluggla, reflekterar fjäderdräkten merparten av UV-strålningen. Dessa fåglar skulle alltså kunna vara "vita" också sedda genom fågelögon, eftersom de reflekterar alla våglängder av det ljus fåglarna ser. Hos andra vita fåglar reflekteras UV-strålning delvis eller mycket lite av fjädrarna. Till dem hör knölsvanen. Forskarna har dock inte testat fjädrar från alla delar av svanens kropp. Knölsvanen är därför troligen inte "vit" sedd genom fågelögon, utan har en färg som vi människor omöjligt kan föreställa oss, åtminstone på delar av kroppen.
Biologiska ytor, oberoende ;av vilken färg de har, är i det närmaste "svartkroppar", när det gäller infraröd strålning av alla våglängder. Det betyder att nästan ingen infraröd strålning reflekteras. Djurs hud absorberar således nästan all infallande infraröd strålning som når den. Detta gäller såväl vit hud, svart hud som hud av alla andra färger. Det samma gäller päls och fjäderdräkt med olika färg, om den inte är så gles att en del infraröd strålning når huden. Nästan all infraröd strålning som når huden absorberas där, oberoende av vilken färg huden har.
När det gäller förluster av värme från kroppen genom strålning, spelar kroppsytans färg ingen roll. Detta beror på att all strålning som avges från hud, fjädrar och päls är infraröd strålning. Vid de relativt låga temperaturer, under vilka liv kan existera, utsänder inga kroppar temperaturorsakad strålning inom det för människan synliga spektralområdet. Solen, med en yttemperatur på cirka 5000 grader Celsius, sänder däremot ut synligt ljus.

Så långt de teoretiska övervägandena. Det finns några undersökningar av den synliga solstrålningens betydelse för vita fåglars värmebalans. I en studie jämförde forskare solbelysta svarta och vita duvor. Vid låga vindstyrkor tog de svarta duvorna upp mycket mer värme i kroppen än de vita, eftersom de reflekterade lite och absorberade mycket av den synliga solstrålningen, utan att den bildade värmen fördes bort genom vindens konvektion. Vid höga vindstyrkor tog både de svarta och de vita duvorna upp betydligt mindre värme än vid låga, eftersom den bildade värmen fördes bort av vinden. Men de vita duvorna tog då upp mer värme än de svarta eftersom en stor del av solstrålningen trängde djupt in i fjäderdräkten och inte blåste bort. Skillnaden var dock ganska liten. I en annan studie jämförde forskare de två färgvarianterna av snögås, den vita och den grå. De kunde inte säkert påvisa någon effekt av färgskillnaden. En studie av olika hägerarter i Floridas varma klimat tydde på att vita hägrar var mindre värmestressade, förmodligen beroende på att deras fjäderdräkt reflekterade en stor andel av den synliga solstrålningen. I en studie av dalripor i stillastående luft fann forskarna att ripor med mörk fjäderdräkt värmdes upp mer av solstrålning än vita ripor. De fann ingen skillnad i uppvärmning mellan mörka och vita ripor, när det blåste.
Slutsatser
I en översiktsartikel gjordes en genomgång av alla föreslagna funktioner för vit fjäderdräkt. Slutsatsen blev att det inte finns starkt experimentellt stöd för någon av hypoteserna. Forskarna föreslog en alternativ hypotes, nämligen att vit färg är ett grundtillstånd och att fåglar blir vita, om färgad fjäderdräkt inte gynnas av det naturliga urvalet. I och för sig sparar vita fåglar på energi genom att de slipper producera pigmentet melanin till fjädrarna. Men denna hypotes synes mig osannolik och den saknar experimentellt stöd. Darwins fråga förblir obesvarad. Hos flertalet vita fåglar vet vi inte vilken funktion, om någon, den vita färgen har.
Fördjupning
- Fåglarnas färgrika fjädrar och däggdjurens hårstrån: hur de bildas och får sin färg. Pigment och strukturfärg.Du hittar artikeln under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?
- Ska jag ha vita eller svarta kläder i stark värme, om jag vill undvika överhettning? Artikeln finns under rubriken Att hålla sig varm när det är kallt, och kall när det är varmt – reglering av djurs kroppstemperatur.
Referenser
- B. A. Beasley och C. D. Ankney: Physiological responses of cold-stressed blue and snow phase lesser snow goose goslings (Canadian Journal of Zoology 70:549-552, 1992).
- D. Burkhardt: UV vision: a bird's eye view of feathers (Journal of Comparative Physiology A 164:787-796, 1989).
- H. I. Ellis: Metabolism and solar radiation in dark and white herons in hot climates (Physiological Zoology 53:358-372,1980).
- W. L. N. Tickell: White Plumage (Journal of the Waterbird Society 26:1-128, 2003).
- J. M. Ward et al.: Thermal consequences of turning white in winter: a comparative study of red grouse Lagopus lagopus scoticus and Scandinavian willow grouse L. l. lagopus (Wildlife Biology 13:120-129, 2007).
- B. O. Wolf och G. E. Walsberg: The role of the plumage in heat transfer processes of birds (American Zoologist 40:575-584, 2000).

Visste du att skallerormar kan "se" utan att använda ögonen och utan att använda synligt ljus? Att de har inbyggda IR-kameror? Däggdjur och fåglar är jämnvarma ("varmblodiga") djur. Detta innebär att de håller en konstant kroppstemperatur, som är högre än omgivningens. Därför sänder de ut en intensivare värmestrålning än omgivningen. Skallerormarna använder sig av denna värmestrålning för att ta reda på var deras byte finns.
Värmestrålning kallas också för infraröd strålning, och de infraröda "ögonen" hos ormar kallas groporgan. Groporgan finns hos en del boaormar (familjen Boidae), de flesta pytonormar (familjen Pythonidae) och hos alla näsgropsormar (underfamiljen Crotalinae bland huggormarna). De mest kända näsgropsormarna är skallerormarna. Hos boaormar och pytonormar finns groporganen intill läpparna, 3-20 organ på varje sida. Näsgropsormarna har ett par groporgan, belägna mellan ögonen och näsborrarrna. I fortsättningen beskrivs bara groporganen hos näsgropsormarna.
Infrarödsinnet hos skallerormarna fungerar inte bara som ett extra sinne, ett "sjätte sinne". Informationen från näsgroparna behandlas nämligen av ormen tillsammans med informationen från ögonens tre typer av tappar. Tapparna är känsliga för olika färger, alltså olika spektralområden av synlig elektromagnetisk strålning. Groporganen är känsliga för ett visst spektralområde av infraröd, för människor osynlig, elektromagnetisk strålning. Informationen om dessa fyra spektralområden når samma områden i hjärnan och integreras där till en bild av omgivningen. Ormarna har alltså ett seende som inkluderar infraröd strålning, precis som fåglar och många insekter har ett seende som inkluderar för oss osynlig ultraviolett strålning. Den ultravioletta strålningen registreras emellertid av en särskild typ av tappar i ögat, inte av ett särskilt organ. Det bör framhållas att groporganet ger en avsevärt sämre bild än ögat.
Hos ryggradsdjur förmedlar det så kallade somatosensoriska nervsystemet sinnesintryck från huden. bland annat av temperatur, av smärta samt av tryck och annan mekanisk stimulering. Trigeminusnerven är en hjärnnerv som leder somatosensorisk information från ansiktet till hjärnan. Hos skallerormar är en del av trigeminusnervens nervfiberändar mycket känsliga termoreceptorer ("temperaturmätare") i groporganen.

Groporganen hos näsgropsormarna utgörs av två små hålor framför och under ögonen. I varje håla finns ett värmekänsligt membran som är cirka 30 kvadratmillimeter stort, men bara 15 µm (0,015 mm) tjockt. Membranet sitter utspänt i hålan och innehåller tusentals ansamlingar av nervfiberändar, som fungerar som termoreceptorer. I nervfiberändarnas cellmembraner finns proteinkanaler, vars öppningsgrad bestäms av deras temperatur. När kanalerna är öppna släpper de in natriumjoner och kalciumjoner i nervfiberändarna, vilket leder till att nervimpulser skickas via nervändarnas nervfibrer i riktning mot hjärnan. Det löper ständigt nervimpulser i fibrerna. När ormen "ser" ett föremål som är varmare än omgivningen, till exempel en mus, värms receptorerna av värmestrålning från musen. Då sänder fibrerna nervimpulser med kortare tidsmellanrum. När ormen "ser" ett föremål som är kallare än omgivningen, till exempel en groda, minskar värmestrålningen, receptorerna kyls och nervimpulser sänds med längre tidsmellanrum.
Nervfiberändarna sitter bara ett par tusendelar av en millimeter från membranytan. Dessutom finns det bakom membranet en ficka fylld med luft som isolerar membranet. Den isolerande luften gör att nästan all värmestrålning absorberas av membranet och påverkar dess temperatur, samt att mycket lite värmestrålning passerar membranet. Den värmestrålning som passerar absorberas av luftfickans bakre vägg. Dessa och andra anpassningar gör receptorerna mycket känsliga för små temperaturförändringar. Det räcker att temperaturen ökar eller minskar med så lite som 0,001 °C för att nervändarna ska reagera.
Groporganets membran har på sin yttre yta en speciell struktur som gör att synligt ljus reflekteras och inte värmer upp receptorerna. Därmed säkerställs att receptorerna bara reagerar på värmestrålning. Membranet innehåller en mängd speciellt utformade blodkapillärer och är väl genomblött. Detta gör att temperaturen hos ett uppvärmt eller nerkylt område snabbt återställs genom att blodet tar upp eller avger värme till området. Funktionen med detta tros vara att hindra den infraröda bilden från att stanna kvar för länge i membranet. I så fall skulle den bilda en efterbild som registreras samtidigt som nästa bild, ungefär som vid en dubbelexponering med en gammaldags kamera.
Själva öppningen på gropen har endast en tredjedel av membranets diameter, vilket gör att groporganet fungerar som en hålkamera. En hålkamera är en linslös kamera, i vilken ett litet hål fungerar som lins. Groporganet producerar alltså i princip en infraröd bild på sitt membran, på samma sätt som ögat producerar en bild av ljus på näthinnan. Upplösningen är dock avsevärt sämre hos groporganet än hos ögat, vilket gör att groporganets bild blir väldigt suddig och oklar. Det område som varje ansamling av nervändar registrerar utgörs av en kon, med spetsen i membranet. En mindre öppning hade gjort denna kon smalare och gett groporganet en högre upplösning, men på bekostnad av ljuskänsligheten. Detta hade förmodligen varit en nackdel.
Du kan själv tillverka en hålkamera genom att sticka ett hål i ett svart papper och sedan försöka projicera den uppochnervända bilden av en lampa eller ett ljust fönster mot den motsatta vita väggen i ett för övrigt mörkt rum.

De områden i hjärnan som är ansvariga för att behandla syninformationen behandlar också informationen från värmestrålningssinnet. Information från ögonen används alltså tillsammans med information från groporganen för att fånga byten. Det finns nervceller som bara reagerar på information från ögonen, andra celler som bara reagerar på information från groporganen och åter andra som reagerar på information från båda dessa sinnesorgan. En typ av nervceller i skallerormshjärnan reagerar maximalt på ett litet, varmt föremål som rör sig. Dessa celler mottar information både från ögonen och från groporganen. De har kallats "musdetektorer". Förmodligen reagerar ormarna främst på på rörliga varma objekt, snarare än på stillastående. Med sina musdetektorer kan ormen skilja mellan en mus och en musliknande sten. Stenen retar inte groporganen, men den varma musen gör det. Det skulle ju inte vara så lyckat om ormen högg mot stenar.
Skallerormar använder sig av luktsinnet vid jakt. De känner också av vibrationer i marken. Men det är synen kombinerad med infrarödsinnet som spelar störst roll när en skallerorm närmar sig sitt byte och utdelar sitt dödande bett. Bettet är mycket snabbt. Ändå hinner ormen med att röra huvudet mot bytet, spänna ut huggtänderna, injicera giftet och dra tillbaka huvudet. Bettet måste vara mycket precist, och det är tack vare information framför allt från ögonen, men även från groporganen, som skallerormar har så bra träffsäkerhet.
Groporganen hjälper sannolikt också skallerormar att reglera sin kroppstemperatur, något som kan ha varit organens ursprungliga funktion. Ormar är växelvarma ("kallblodiga") djur, som inte producerar sin egen kroppsvärme. De kan höja sin kroppstemperatur bland annat genom att uppsöka varma områden i sin omgivning. Groporganen kan hjälpa skallerormar att hitta sådana områden. Märkligt nog tycks skallerormar inte använda sina groporgan för att orientera sig i mörker.
Referenser
- R. C. Goris: Infrared organs of snakes: an integral part of vision (Journal of Herpetology 45:2-14, 2011).
- K. V. Kardong och H. Berkhoudt: Rattlesnake hunting behavior - correlations between plasticity of predatory performance and neuroanatomy (Brain, Behavior and Evolution 53:20-28, 1999).
- A. R. Krochmal, G. S. Bakken, och T. J. LaDuc: Heat in evolution’s kitchen, evolutionary perspectives on the functions and origin of the facial pit of pitvipers (Viperidae: Crotalinae) (Journal of Experimental Biology 207:4231-4238, 2004).
- E. A. Newman och P. H. Hartline: The infrared "vision" of snakes (Scientific American 246:98-107, 1982).
- H. A. Schraft, G. S. Bakken, och R. W. Clark: Infrared-sensing snakes select ambush orientation based on thermal backgrounds (Scientific Reports 9:3950, 2019).
- H. A. Schraft och R. W. Clark: Sensory basis of navigation in snakes, the relative importance of eyes and pit organs (Animal Behaviour 147:77-82, 2019).

Hur kan ett rovdjur som inte använder syn och hörsel ändå effektivt fånga sitt byte, på natten och under vattnet dessutom? För näbbdjuret (Ornithorhyncus anatinus) har gåtan ett svar långt mer spännande än någon tidigare vågat tro. Näbbdjuret har receptorer, mottagare av sinnesstimuli, på näbben som registrerar de elektriska fält som alstras av bytesdjuren. Dessa elektriska fält är mycket svaga. Tester har visat att näbbdjuret kan uppfatta mycket låga fältstyrkor (cirka 0,000 020 V/cm). Med hjälp av elektroreceptorer och mekanoreceptorer på näbbens ytor kan näbbdjuret bedöma riktningen och kanske också avståndet till ett bytesdjur. Mekanoreceptorerna mäter vågrörelser i vattnet.
Näbbdjuret är ett däggdjur och räknas till gruppen kloakdjur (Monotremata). Kroppslängden är omkring 0,5 meter och vikten ungefär 1-2 kg. Honan lägger ägg och efter kläckningen ger hon ungen di. Näbbdjuret lever i och vid sötvattenssystem i östra Australien och på Tasmanien. Det är ett nattaktivt rovdjur som livnär sig på sötvattenslevande ryggradslösa djur, till exempel kräftdjur och insektslarver. Djuret söker föda i vattnet med hjälp av sin känsliga näbb, som det för fram och tillbaka över bottnen. Det vänder även på stenar och bökar i bottenmaterialet.
Näbbdjuret dyker med ögon, öron och näsöppningar stängda. Med stora simhudsförsedda framfötter och en tät päls är den väl anpassad för ett vattenliv. Det svaga elektriska fältet från ett gömt eller flyende bytesdjur utlöser en automatisk liten vridning av huvudet i riktning mot bytesdjuret. Bytesdjuren alstrar även vågor i vattnet som fortplantas långsammare än de elektriska strömmarna. Näbbdjuret reagerar också på sådana vågor.
Det finns cirka 100 000 elektroreceptorer och mekanoreceptorer på insidan och utsidan av näbben, ungefär hälften av varje sort. En elektroreceptor består av en nedsänkt por i näbbens hud. I porens vägg finns ett flertal nervfiberändar, som sticker in i hålrummet. Det är dessa fiberändar som känner av elektriska fält. Nervfibrerna leder signaler till hjärnan.
Näbbdjuret skapar förmodligen med hjälp av det stora antalet elektroreceptorer på näbben en karta över det elektriska fältet i omgivningen. Med hjälp av kartan kan djuret sedan bedöma var fältet har sitt ursprung. Med mekanoreceptorerna känner den samtidigt av när mekaniska vattenrörelser når näbben. Vattenrörelserna är långsammare än de elektriska strömmarna. Skillnaden i tid mellan den elektriska och den mekaniska stimuleringen känns av i så kallade bimodala nervceller och ger förmodligen information om avstånd. Med hjälp av sin känsliga näbb kan näbbdjuret alltså bedöma inte bara riktningen, utan troligen också avståndet, till ett bytesdjur.
Signalerna från receptorerna sammanställs i hjärnan. I en viss del av storhjärnsbarken, ;S1, ligger skivor med nervceller som förmedlar signaler från elektroreceptorer omväxlande med skivor som förmedlar signaler från mekanoreceptorer. Skivorna bildar alltså en randig struktur. I de skivor som tar emot information från elektroreceptorerna finns också nervceller av bimodal typ, det vill säga celler som reagerar både på elektroreceptorsignaler och mekanoreceptorsignaler. Det är troligen sådana celler som kan ge hjärnan information om avståndet till bytet. S1-delen av näbbdjurets hjärna liknar faktiskt delar av vår storhjärnsbark som bland annat tros ha en viktig funktion för avståndsbedömningen vid seendet.
Det var så nyligen som på 1950-talet som forskare uppmärksammade djurs förmåga att känna av svaga elektriska fält. Denna förmåga finns både hos broskfiskar (hajar och rockor) och hos flera grupper av benfiskar. På 1980-talet uppdagades egenskapen även hos näbbdjuret och dess släktingar, myrpiggsvinen. Det var mycket oväntat att hitta elektroreceptorer även hos däggdjur.
Luft är en isolator som inte leder elektriska strömmar. Det må då synas märkligt att de helt landlevande myrpiggsvinen har ett elektriskt sinne. Det kan bara komma till användning i väta, till exempel när myrpiggsvinen letar efter föda med näbben i våt jord. Myrpiggsvinen har emellertid ett mycket sämre elektriska sinne än näbbdjuret. Förmodligen härstammar de från vattenlevande kloakdjur med välutvecklat elektrisk sinne och är på väg att förlora de, nu när de övergått till landliv.
En mycket viktig sak som näbbdjursforskningen bidragit till är införandet av störningsfria inhägnader åt näbbdjuren i djurparker. De skärmar helt enkelt av alla elektriska störningar med en så kallad Faradays bur. Den behöver inte vara en bur i ordets vanliga bemärkelse, utan den kan bestå av nät av tunna jordade elledare som fångar upp störande elektriska fält från elektrisk utrustning i djurparken, innan de når näbbdjuren.
Referenser
- Australian Platypus Conservancy (nedladdat den 4 juni 2013)
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012)
- J.D. Pettigrew: Electroreception in monotremes (Journal of Experimental Biology 202:1447-1454, 1999)
Att hålla sig varm när det är kallt, och kall när det är varmt – reglering av djurs kroppstemperatur

Människan är jämnvarm (homeoterm, "varmblodig"), i likhet med nästan alla andra däggdjur och fåglar. Det innebär att hon håller kroppstemperaturen nära nog konstant dygnet runt. Hon reglerar kroppstemperaturen genom flera intressanta mekanismer.
Vi brukar säga att människans normala kroppstemperatur är cirka 37,0±0,5 °C. Det finns dock förvånande stora individuella skillnader i kroppstemperatur, också mellan friska personer. Däggdjur och fåglar har också olika kroppstemperaturer.
Kroppstemperaturen varierar inom vissa gränser
Hos alla människor uppvisar kroppens temperatur en dygnsrytm (cirkadiansk rytm) med en spännvidd på cirka 0,5-0,8 °C. Den är lägst sent under natten och högst under sen eftermiddag eller tidig kväll. Kroppstemperaturen påverkas även av en rad andra faktorer. Under loppet av kvinnors menstruationscykel stiger den något efter ägglossningen. Vid intensivt muskelarbete kan den normalt stiga till uppemot 40 °C. Denna temperaturhöjning anses, åtminstone delvis, bero på att kroppens värmeproduktion är så hög, att vi inte klarar av att bibehålla kroppstemperaturen. I varma och fuktiga miljöer kan temperaturen stiga ytterligare, något som kan leda till värmeslag i samband med arbete. Feber är märkligt nog en kontrollerad höjning av den temperatur som kroppens termostat är inställd på.
Temperaturen regleras bara i kroppskärnan. Värmeväxlare finns i extremiteterna
Temperaturen är faktiskt inte strikt reglerad i hela kroppen. Det är bara temperaturen i kroppens kärna som hålls inom snäva gränser. Kärnan är de inre delarna av huvudet och bålen. I huden och inuti armar och ben varierar temperaturen avsevärt mer. Den är där oftast lägre än i kroppskärnan. Temperaturen i testiklarna hålls låg, vid cirka 32 °C. Detta sker genom att artärerna till dem och venerna från dem bildar värmeväxlare, som håller kvar värme i bålen. Detta är nödvändigt för att spermieproduktionen ska kunna ske. Värmeväxlare finns också i armar och ben. De håller dessa extremiteter kallare än normalt i kalla miljöer.

Vad krävs för att hålla kroppstemperaturen konstant? Hur transporteras värme?
Låt oss då förklara hur vi reglerar kroppstemperaturen. Vi förutsätter att vi är i vila, vilket innebär att vi inte rör på oss, och att omgivningens temperatur är lägre än kroppens temperatur. Vi förutsätter också att vi befinner oss i skugga, så att vi inte tar upp värme från omgivningen, framför allt solstrålning. För att hålla en konstant kroppstemperatur måste vi då producera precis lika mycket värme som vi avger till omgivningen. Jämnvarma djur har en stor förmåga att själva producera värme i kroppen. Detta är en nödvändig förutsättning för att de ska kunna hålla sin kroppstemperatur nära nog konstant. Vi avger värme till miljön genom värmestrålning (IR-strålning), genom ledning av värme och genom så kallad konvektion. Ledning sker huvudsakligen genom att molekyler påverkar varandra, i synnerhet om kroppen är i kontakt med ett fast föremål eller en vätska. Vi har alla upplevt hur vi kyls, när vi sitter på ett kallt underlag eller när vi simmar i kallt vatten. Konvektion sker genom att luften intill oss rör på sig. Vi har alla upplevt hur vi svalkas av en sommarvind och hur vi kyls ner av en vinterstorm.
Termoneutral zon: det "behagliga" temperaturområdet i miljön
Den vilande kroppens ämnesomsättning är som lägst och konstant inom ett yttertemperaturområde som kallas den termoneutrala zonen eller TNZ. Den termoneutrala zonen för en naken fastande människa ligger vanligen mellan cirka +27–28 °C och +31–32 °C i vindstilla luft och skugga. En naken människa upplever dessa omgivningstemperaturer som behagliga. Det finns dock individuella variationer, sannolikt också skillnader mellan olika folkgrupper. Dessutom ändras TNZ förmodligen, när du acklimatiserar sig till höga eller låga yttertemperaturer. Ibland anger vi temperaturkomfortzonen, det vill säga det som människor subjektivt upplever som det behagliga temperaturområdet. Det uppges ligga på ungefär samma nivåer som TNZ, men med ett smalare temperaturspann. TNZ sjunker till lägre temperaturer om du isolerar dig med kläder.
Vår termoneutrala zon ligger högt på temperaturskalan. Detta beror på att människan fortfarande är ett tropiskt djur. Vi har ju till och med förlorat den värmeisolerande pälsen. Det är endast tack vare kläder som vi kunnat sprida oss till kallare områden på jorden.
Reglering i kall miljö
Vid yttertemperaturer under den termoneutrala zonen ökar våra värmeförluster och kroppens temperatur sjunker något. Vi försvarar då kroppstemperaturen genom att huttra. Huttrande är ofrivilliga lokala sammandragningar i skelettmusklerna. Vi märker av dem när vi hackar tänder. Muskelarbetet vid huttring leder till att ämnesomsättningen ökar. Då produceras mer spillvärme som används till att återställa kroppstemperaturen.
Människan och andra däggdjur kan också öka värmeproduktionen i kroppen utan hjälp av muskelsammandragningar. Detta kallas "non-shivering thermogenesis", vilket kan översättas med icke-huttrande värmeproduktion. Många däggdjur, i synnerhet på nordliga breddgrader, huttrar inte alls eller sällan.
Hos många däggdjur, särskilt små sådana, äger den icke-huttrande värmeproduktionen rum i det bruna fettet, ett slags biologiskt värmeelement. Forskarna har länge vetat att spädbarn använder brunt fett för att värma upp sig. Det bruna fettets roll hos vuxna människor har dock varit oklar. Forskare har emellertid nu påvisat en ökad värmeproduktion i det bruna fettet vid kyla även hos vuxna människor. Dessutom har de funnit att icke-huttrande värmeproduktion även sker i vanligt fett, vit fettvävnad, både hos spädbarn och vuxna. Där sker det i så kallade beigefärgade fettceller. Det finns också studier som antyder att brunt fett kan bidra till att reglera kroppsvikten hos människor genom att aktiveras vid ökat födointag. Därmed skulle det bidra till att förhindra en viktuppgång.
Icke nog med detta, forskare har på senare tid hos människan och andra däggdjur påvisat icke-huttrande värmeproduktion i skelettmuskelceller, alltså en ökad värmeproduktion i dessa celler utan att de drar ihop sig. Det handlar om så kallade futila cykler, ett ökat läckage av joner ut ur de menbranomslutna cellkomponenter som kallas mitokondrier och endoplasmatiskt nätverk. Dessa joner pumpas ständigt in i dessa cellkomponenter. Pumpningen är energikrävande. Om läckaget ökar går pumparna på "tomgång" och värmeproduktionen ökar.

Reglering i varm miljö
Vid yttertemperaturer ovanför den termoneutrala zonen minskar våra värmeförluster och kroppstemperaturen stiger. Om vi lämnar skuggan, tar vi till och med upp värme från den brännande solen. I båda fallen måste vi försvara kroppens temperatur genom avdunstning av vatten (evaporation). När vatten avdunstar från kroppen, överförs värme från kroppen till vattenmolekylerna i det gasformiga vatten (vattenånga) som bildas. Vi människor kyler oss genom att svettas och låta svetten avdunsta. Även då ökar ämnesomsättningen, eftersom det krävs energi för att producera svett. Den mängd spillvärme, som då produceras i kroppen, är emellertid betydligt mindre, än den mängd värme, som vi gör oss av med när svetten avdunstar. De flesta däggdjur flämtar i stället för att svettas.
Om det finns vattensamlingar eller vattendrag i närheten, kan däggdjur och fåglar även kyla sig genom att bada. När de stiger upp ur badet, är kroppen täckt av vatten. När detta vatten avdunstar, åtgår värme och huden kyls. Detta fungerar bäst om pälsen eller fjäderdräkten är tunn. Allra bäst fungerar det hos däggdjur som saknar päls, till exempel människor och elefanter. Fördelen med denna kylmekanism är att djuren inte förlorar dyrbart kropsvatten i en varm och torr miljö, men den kräver naturligtvis tillgång till vatten någonstans i miljön.
Elefanter har en skrynklig hud. Afrikanska elefanter har dessutom rikligt med mycket tunna sprickor i huden, cirka 0,1 mm breda. Efter ett bad kan vatten stanna kvar länge i skrynklorna och sprickorna. Därmed kan elefanten kylas genom avdunstning långt efter den har badat.
Reglering i "behaglig" miljö
Vid yttertemperaturer inom den termoneutrala zonen kan vi reglera kroppstemperaturen genom att ändra blodflödet till huden och därmed kroppens värmeisolering. Är vi varma så ökar vi blodflödet och därmed värmeavgivningen. Huden blir röd. Fryser vi så minskar vi blodflödet och därmed värmeavgivningen. Huden blir vit. När de flesta andra däggdjur fryser, kan de dessutom öka sin värmeisolering genom att höja pälsens hår, "resa ragg". En tjockare päls minskar värmeförlusterna. När dessa djur är varma, sänker de pälsen och minskar då sin värmeisolering. Vi människor kan också höja och sänka pälshåren. Men eftersom de flesta håren är så små att de inte syns påverkar vi inte vår värmeisolering. När vi fryser yttrar sig raggresningen bara som så kallad gåshud. Men vi kan ersätta pälsen med kläder. Då kan vi öka och minska vår värmeisolering genom att anpassa klädedräkten efter vädret.
Vad händer i mycket stark hetta?
Men vad händer om vi befinner oss i en miljö med en högre temperatur än vi själva eller om vi är utsatta för mycket intensiv solstrålning, till exempel under en lång ökenvandring? Då tar vi upp mera värme, än vi klarar av att avge till omgivningen. Nu finns det bara ett sätt att försvara kroppens temperatur: genom avdunstning av svett. Vattenförlusten bli då mycket stor. Du kan avge flera liter svett i timmen. Så länge vi har god tillgång till dricksvatten finns det hopp om livet. Vi måste dock skydda huvudet så att hjärnan inte överhettas. Det leder till värmeslag (solsting). Men när dricksvattnet tagit slut, klarar vi oss inte länge. Vattenförlusten leder snart till blodtrycksfall och sedan till cirkulatorisk chock. Utan medicinsk hjälp överlever vi då inte. Men vi får hoppas att vi dessförinnan upptäcks av räddningsmanskapet.
Referenser
- N.C. Bal et al: Sarcolipin is a newly identified regulator of muscle-based thermogenesis in mammals (Nature Medicine 10:1575-1579, 2012).
- K.E. Barnett, S.M. Barman, S. Boitano, och H.L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- S.D. Clarke et al: Postprandial heat production in skeletal muscle is associated with altered mitochondrial function and altered futile calcium cycling. (American Journal of Physiology - Regulatory, Integrative and Comparative Physiology 303:R1071–R1079, 2012).
- A.M. Cypess et al: Anatomical localization, gene expression profiling and functional characterization of adult human neck brown fat (Nature Medicine 19:635-9, 2013).
- H. Færevik, D. Markussen, G.E. Øglænd och R.E. Reinertsen: The thermoneutral zone when wearing aircrew protective clothing (Journal of Thermal Biology 26:419-425, 2001).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- P. Willmer, G. Stone, och I. Johnston: Environmental Physiology of Animals (2nd ed, Blackwell, 2005).

Mellan vilka kroppstemperaturer kan djur och andra organismer överleva? Den nedre gränsen sätts för många organismer av att de fryser till is vid en temperatur strax under 0 °C. Köldkänsliga organismer kan dö vid betydligt högre temperaturer. Den övre gränsen sätts för många organismer av att proteiner och andra stora molekyler förändras så att de slutar att fungera, ofta vi 40–50 °C, för värmekänsligare organismer vid betydligt lägre temperaturer.
Det finns emellertid många organismer, som kan vara aktiva utanför temperaturspannet från ungefär 0 till ungefär 40–50 °C. Det finns också många organismer, som kan klara mycket extremare temperaturer genom att gå in i ett vilstadium. Naturligtvis är levande varelser särskilt köldtåliga i tempererade och arktiska klimat, särskilt värmetåliga i tropiska klimat.
Fiskar i kalla hav: antifrysmedel och underkylning
Fiskar i de arktiska och antarktiska haven lever i en omgivning med en ständig temperatur på ca -2 °C. Havsvattnets fryspunkt är nämligen lägre än rent vattens på grund av att lösta ämnen, främst natriumjoner och kloridjoner, sänker dess fryspunkt. Dessa fiskars kroppstemperatur är lika med vattnets, alltså cirka -2 °C. De flesta av dem har en mycket lägre koncentration av lösta ämnen inuti sig än havsvattnets och skulle alltså frysa till is, om de inte hade speciella försvarsmekanismer. En del av dem har i sina kroppsvätskor speciella glykoproteiner som sänker fiskens fryspunkt genom att binda till nybildade iskristaller och hindra dem från att växa. Andra fiskar är underkylda.

Insekter på vintern: lösta ämnen sänker fryspunkten
Många insekter och andra ryggradslösa djur på land överlever vintern i ett vilande tillstånd. De övervintrande djuren kan använda båda de mekanismer, som beskrivs för fiskar ovan, för att undgå att frysa till is. De kan också sänka sin fryspunkt genom att de skaffar sig höga koncentrationer av sockerarter eller sockeralkoholer i kroppsvätskorna. Dessa substanser är "proteinvänliga" och kan förkomma i hög koncentration utan att denaturera proteiner. Slutligen kan en del av dessa djur tåla isbildning i vävnaderna. Vissa av dem kan i vilstadium överleva åtminstone -20 °C. Några insekter och spindlar kan till och med vara aktiva vid frysgrader.
Djur som tål att frysa till is
Vissa djur är frystoleranta och tål att frysa till is, förutom vissa insekter, även många djur som lever på nordliga tidvattensstränder. De har då "groddar" utanför cellerna som gör att isen bildas i vätskan där och inte inne i cellerna. Cellerna undgår därför att frysa till is, något som skulle vara fatalt. Det finns till och med nordligt levande grodor och ormar, som kan överleva isbildning i kroppen.
Det finns inga organismer som kan vara aktiva och föröka sig infrusna i is. Men det finns organismer, till exempel bakterier, som kan leva sitt liv i vattenfickor inuti isen.

De fantastiska björndjuren
Bland djuren är trögkryparna (björndjuren) särskilt kända för att tåla extremt låga temperaturer. De gör detta i ett vilstadium, så kallad kryptobios. De är de köldtåligaste och värmetåligaste djur vi känner till. I kryptobios överlever de från -272 till +150 °C. De kan dock inte vara aktiva vid de extrema temperaturerna.
Däggdjur och fåglar i kallt klimat
Däggdjur och fåglar kan hålla kroppstemperaturen konstant i kalla miljöer. Det gör de genom att själva producera värme i kroppen och genom att vara väl isolerade mot kylan med tjock päls eller fjäderdräkt. Det är ingen tillfällighet att dessa två grupper är de enda landryggradsdjur som kan leva i Arktis och Antarktis. Där kan flera av dem tåla lufttemperaturer på -30 °C eller lägre utan att skadas.

Extremt värmetåliga ryggradslösa djur
När det gäller den övre temperaturgränsen, så finns det organismer utrustade med proteiner och andra cellbeståndsdelar som är mycket tåliga mot värmedenaturering. Det finns ryggradslösa djur som klarar av kroppstemperaturer på upp emot 50–55 °C. Till dem hör pompejimasken (Alvinella pompejana), musselkräftan Potamocypris och ökenlevande myror av släktet Cataglyphis.
Däggdjur och fåglar i extrem värme
De flesta fåglar håller en kroppstemperatur på cirka 39-42 °C, men i stark hetta kan många fåglar ha kropptemperaturer på 43-46 °C. Vissa stora ökenlevande däggdjur låter kroppstemperaturen stiga under dagen, men sänker den under natten. På dagen kan de ha kropptemperaturer på 45-47 °C. Dessa fåglar och däggdjur höjer kroppstemperaturen under dagen för att slippa kyla sig genom att avdunsta vatten, en sällsynt vara i de miljöer de lever i. De klarar emellertid inte av att hålla dessa höga temperaturer dygnet runt.

Bakterier och arkéer: rekord i värmetålighet
De mest extrema exemplen på att aktivitet vid höga temperaturer hittar vi hos bakterier och arkéer, som lever och förökar sig vid mycket heta källor på land och i djuphavet. Dessa organismer kallas termofiler. Om de klarar av extremt höga temperaturer kallas de hypertermofiler. Landlevande arter klarar av temperaturer från 60 °C till över 90 °C. I havet har forskare funnit arkéer som klarar av cirka 120 °C. Rekordet hålls för närvarande av Methanopyrus kandleri som kan leva och föröka sig vid 122 °C. Termofilerna är mycket intressanta, eftersom de lever i miljöer som kan ha förekommit under den period för cirka 4 miljarder år sedan, då livet tros ha uppkommit på vår jord.
Du undrar kanske hur organismer kan leva vid 120 °C, när vatten kokar vid 100 °C. Men denna kokpunkt gäller vid havsytans nivå. Vattnets kokpunkt stiger då trycket ökar och sjunker då trycket minskar. På 1 000 meters djup under havsytan är trycket cirka 100 gånger högre än vid havsytans nivå. Vattnets kokpunkt är därför 310 °C. På 6 000 meters djup är kokpunkten 480 °C. De värmetåliga arkéerna lever vid skorstensliknande så kallade hydrotermala källor på flera tusen meters djup. Ur dessa källor flödar vatten med en temperatur på flera hundra grader Celsius. På grund av det höga trycket kokar inte vattnet.
Hur celler skyddar sig mot skadliga temperaturer
Växelvarma ("kallblodiga") djur håller i stort sett samma temperatur som omgivningen, Växelvarma djur, som lever vid extrema temperaturer, är utrustade med enzymer och andra proteiner med en struktur, som gör att de fungerar i stark kyla eller i stark värme. Cellernas membraner måste också vara anpassade till temperaturen, annars blir de alltför stela i kyla och alltför "sladdriga" i värme. Membranerna innehåller fettämnen som kallas fosfolipider. Djur som lever i kalla miljöer har en hög halt av omättade fettsyror i sina fosfolipider. Därmed blir membranerna lagom stela vid låg temperatur. Djur som lever i varma miljöer har en hög halt av mättade fettsyror i fosfolipiderna, vilket gör membranerna lagom stela vid hög temperatur.
Alla organismer har skyddsmekanismer, som skyddar dem mot skadliga temperaturer. Så kallade värmechocksproteiner skyddar celler mot extrem värme. De skyddar också mot annan stress, ibland till och med mot kyla. De borde därför kallas stressproteiner. De flesta av dessa proteiner produceras inuti cellerna bara vid stress. De fungerar som så kallade chaperoner. Chaperonerna är cellernas sjukvårdare, som tar hand om skadade, alltså denaturerade, proteiner. De ser till att skadade proteiner återgår till sin normala veckade struktur. Proteiner kan emellertid bli så skadade att de är bortom all räddning. Sådana proteiner måste avlägsnas, innan de skadar cellerna. Detta sker genom att det lilla proteinet ubikvitin binds till dem. De är nu märkta som avfall och tas om hand av proteasomer, proteinkomplex som tillsammans med fria enzymer bryter ner dem till aminosyror.
Fördjupning
Reglering av kroppstemperaturen i värme och i kyla: huttrande, blodflöde, gåshud och svettning. Artikeln finns under rubriken Att hålla sig varm när det är kallt, och kall när det är varmt – reglering av djurs kroppstemperatur.
Referenser
- W. J. Gehring och R. Wehner: Heat shock protein synthesis and thermotolerance in Cataglyphis, an ant from the Sahara desert (Proceedings of the National Academy of Science USA 92:2994-2998, 1995).
- H. Heasler och C. Jaworowski: Life in extreme heat (In: Yellowstone Resources and Issues Handbook 2014, hämtad 9 juni 2015).
- R. W. Hill, G .A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- J. Ravaux et al.: Thermal limit for metazoan life in question, in vivo heat tolerance of the Pompeii worm (PLoS ONE 8: e64074. doi:10.1371/journal.pone.0064074).
- K. Takai et al.: Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation (Proceedings of the National Academy of Science USA 105:10949-10954, 2008).
- Y. Voituron et al.: Freeze tolerance evolution among anurans, frequency and timing of appearance (Cryobiology 58:241-247, 2009).
- C. E. Wickstrom och R. W. Castenholz: Thermophilic ostracod, aquatic metazoan with the highest known temperature tolerance (Science 181:1063-1064, 1973).

Förr trodde vi att dinosaurier var tröga och långsamma djur med låg energiomsättning, i motsats till dem, som vi trodde, "högre" utvecklade däggdjuren. Men vi hade fel. Dinosaurierna dominerade trots allt bland de större landlevande djuren under betydligt längre tid än vi däggdjur har gjort det. Och de är inte utdöda. Fåglarna är nu levande dinosaurier. Dessutom var de utdöda dinosaurierna sannnolikt inte "kallblodiga", alltså växelvarma.
Växelvarma och jämnvarma djur
Forskarna trodde således förr att de utdöda dinosaurierna, precis som de flesta av dagens reptiler, var poikiloterma (växelvarma, "kallblodiga"), det vill säga höll samma temperatur som omgivningen, och att de var ektoterma, det vill säga inte kunde värma upp sig med egenproducerad kroppsvärme. Forskarna trodde också att de inte kunde reglera sin kroppstemperatur, förutom genom beteende, till exempel genom att värma upp sig med hjälp av solvärme.
Nästan alla däggdjur och fåglar är homeoterma (jämnvarma, "varmblodiga"), det vill säga håller en nära nog konstant kroppstemperatur, och endoterma, det vill säga kan värma upp sig med egenproducerad kroppsvärme. De kan också reglera sin kroppstemperatur på en nära nog konstant nivå via andra mekanismer än beteende.
Var dinosaurierna jämnvarma?
Numera har forskarna drastiskt ändrat sin uppfattning om de utdöda dinosaurierna. De flesta menar att de kunde värma upp sig själva och troligen också reglera sin kroppstemperatur. Men hade de en lika hög inre värmeproduktion och en lika effektiv temperaturreglering som däggdjur och fåglar? Här går åsikterna isär. Det handlar ju om fysiologiska egenskaper och sådana fossileras inte. Dessutom har vi mycket få fossil med bevarade mjukdelar. Vanligen har vi bara skelettdelar. Vilka bevis tyder då på att de utdöda dinosaurierna var homeoterma, endoterma reglerare?
Forskarna vet nu att fåglarna är en gren på dinosauriernas släktträd. Vi kan därför misstänka att de andra dinosaurierna hade många egenskaper gemensamma med fåglarna. Men detta är inte hårda fakta.

Hög tillväxthastighet
De fossila dinosaurieägg forskare hittat är förvånande små i förhållande till djurens storlek. Däremot var de nykläckta ungarnas tillväxthastighet förvånande hög. Forskarna har kunnat mäta tillväxthastigheten genom att räkna årsringar i skelettbenen och uppskatta djurens kroppsvikt utifrån benens storlek. Alla undersökta utdöda dinosaurier av olika storlek växte betydligt snabbare än nutida kräldjur. Stora dinosaurier hade tillväxthastigheter i samma storleksordning som nutida däggdjur och fåglar. Denna snabba tillväxt tyder på att de växande dinosaurierna hade en hög energiomsättning och därmed en hög inre värmeproduktion, alltså endotermi.
Stora djur kan lättare hålla sig varma
De stora dinosaurierna var så stora att de sannolikt kunde höja sin kroppstemperatur över omgivningens temperatur, även med en förhållandevis liten värmeproduktion. Detta beror på att större djur har en mindre värmeavgivande kroppsyta mätt per kilo kroppsvikt. Fenomenet kallas gigantotermi. Forskare har påvisat gigantotermi hos några nu levande stora reptiler, nämligen hos stora krokodiler och hos havslädersköldpaddan. Havslädersköldpaddan kan också till en viss del reglera sin kroppstemperatur, åtminstone när den simmar.
Riskerade stora dinosaurier överhettning?
Till detta kommer att klimatet under dinosauriernas tidsålder var varmare än nu. Det var kanske till och med så att mycket stora dinosaurier hade blivit överhettade, om de hade samma höga energiomsättning som dagens däggdjur och fåglar. Nutida elefanter måste ha en rad anpassningar som motverkar överhettning. Och de är betydligt mindre än de största dinosaurierna. Kanske var det så att dinosauriernas snabbväxande ungar hade en hög energiomsättning, men att mycket stora vuxna dinosaurier måste ha haft en betydligt lägre omsättning än däggdjur och fåglar.
Tecken på hög kroppstemperatur
Uppskattningar av dinosauriernas kroppstemperaturer tyder på att de var endoterma, men ger olika resultat. I en studie mätte forskarna isotoper av kol och syre i fossila tänder från stora dinosaurier. Denna studie tyder på att dessa djur hade en kroppstemperatur på 36–38 °C, ungefär som dagens däggdjur. I en annan studie, baserad på tillväxthastighet, kom forskarna fram till en kroppstemperatur på 25 °C för en dinosaurie på 12 kg och 41 °C för en dinosaurie på 13 ton. Forskarna har också gjort fossilfynd, där många individer av flera olika dinosauriearter har hittats tillsammans. I dessa fynd tycks andelen köttätande relativt växtätande individer vara närmare den som gäller för nutida däggdjur än den som gäller för nutida kräldjur. Detta tyder på att de utdöda dinosaurierna hade en hög energiomsättning och därmed en hög inre värmeproduktion, med andra ord att de var endoterma. Dessutom fanns det vissa dinosaurier i polartrakterna, där inga av dagens reptiler kan överleva. Dessa dinosaurier hade knappast kunnat överleva där utan att hålla kroppens temperatur högre än omgivningens. De hade förmodligen också en isolerande fjäderdräkt.

Dinosauriernas hjärta och lungor
Forskarna har länge frågat sig om de utdöda dinosaurierna hade ett trerummigt hjärta, som dagens kräldjur, eller ett fyrrummigt, som dagens däggdjur och fåglar. Problemet troddes vara löst, när de hittade ett bevarat fossilt hjärta. Detta beskrevs som fyrrummigt med en enda aortabåge, precis som däggdjurens och fåglarnas. Fyndet har dock sedermera starkt ifrågasatts. Icke desto mindre tror dock de flesta att de utdöda dinosaurierna hade ett fyrrummigt fågelliknande hjärta. Ett skäl är att långhalsade dinosaurier inte hade kunnat producera ett tillräckligt högt blodtryck för att förse hjärnan med syrerikt blod, om de hade haft ett trerummigt hjärta.
Många skelettben hos utdöda dinosaurier hade stora inre hålrum och var därmed förvånansvärt lätta. Hålrummens struktur tyder på att de var fyllda med luft, precis som hos fåglar. Hos fåglarna är de luftfyllda hålrummen delar av de så kallade luftsäckarna. Luftsäckarna ingår i fåglarnas andningsapparat, som ger dem ett avsevärt effektivare syreupptag än däggdjurens. Revbenens struktur hos flera utdöda dinosaurier tyder på att de hade en fågelliknande andningsmekanism. Allt detta stödjer tanken att de utdöda dinosaurierna hade en liknande andningsapparat som fåglarna. De kan därför ha haft ett stort syreupptag, vilket möjliggjorde en hög energiomsättning och värmeproduktion, alltså endotermi.
Om hur lungornas funktion och globala katastrofer kan ha påverkat dinosauriernas och däggdjurens evolution
Under karbonperioden och början av permperioden var syrehalten i atmosfären upp emot 30 %, mycket högre än dagens 21 %. Med början under perm för cirka 300 miljoner år sedan sjönk sedan halten för att under triasperioden (cirka 250–200 miljoner år före nutid) nå en bottennivå på cirka 10 %. Vid övergången mellan perm och trias ledde dessutom globala katastrofer till det största massutdöendet av djurarter någonsin. De dominerande större landdjuren under perm var så kallade synapsida reptiler, bland dem föregångare till dagens däggdjur. De flesta av dem dog ut.
Under trias skedde i stället en snabb utveckling av dinosaurier. De fyllde de lediga ekologiska nischerna och blev nu de dominerande större landdjuren. Forskarna tror att detta åtminstone delvis berodde på att dinosaurierna hade en fågelliknande andningsapparat och därmed lättare klarade av luftens låga syrehalt. En del synapsider klarade sig dock och gav så småningom upphov till däggdjuren. Men detta skedde först efter det stora utdöendet vid meteoritnedslaget för cirka 70 miljoner år sedan. Då ökade däggdjuren kraftigt i antal. Eftersom alla dinosaurier utom fåglarna hade dött ut, kunde däggdjuren ta över en mängd tomma nischer. Syrehalten i luften låg då på ungefär samma nivå som nu.
Inte växelvarma, inte jämnvarma, men mitt emellan?
Hos nu levande djur finns det ett matematiskt samband mellan tillväxthastighet och energiomsättning i vila. I en studie från 2014 använde forskare detta samband till att uppskatta energiomsättningen i vila hos flera utdöda dinosaurier. De uppskattade också djurens kroppsvikt utifrån skelettbenens dimensioner. Sedan konstruerade de ett diagram som jämförde energiomsättning och kroppsvikt hos de utdöda dinosaurierna med ett stort antal nutida djur, både jämnvarma och växelvarma.
De utdöda dinosaurierna hamnade i diagrammet mellan de jämnvarma och de växelvarma djuren. Forskarna drog slutsatsen att dinosaurierna hade en medelhög energiomsättning och att deras kroppstemperatur var högre än omgivningens temperatur, men inte lika hög som hos nutida jämnvarma däggdjur och fåglar. Forskarna antog att de hade en varierande kroppstemperatur, som reglerades inom vida gränser eller inte reglerades alls. Detta mellanläge mellan endotermi och ektotermi kallade de mesotermi. Vissa nutida djur klassificerades också som mesoterma, bland annat vissa fiskar, den ovan nämnda havslädersköldpaddan samt, bland däggdjuren, kloakdjuren. Väger vi samman den information som finns, förefaller detta vara en tilltalande hypotes. Å andra sidan borde det rimligen ha funnits stora skillnader i temperaturanpassning mellan olika grupper av dinosaurier.
Var fisködlor och flygödlor också jämnvarma?
Det finns data som tyder på att inte bara dinosaurierna, utan också de utdöda fisködlorna (ichthyosaurierna) och flygödlorna (pterosaurierna), var jämnvarma. Dessa grupper var inga ödlor och inte heller dinosaurier. De var kräldjur evolutionärt anpassade till simning respektive flygning och var inte nära släkt med andra reptilgrupper. De var samtida med dinosaurierna.
Fisködlorna är ett klassiskt exempel på så kallad konvergent evolution, det vill säga oberoende utveckling av liknande anpassningar hos obesläktade djur med likartat levnadssätt. De påminde mycket om delfiner och var liksom de senare marina snabbsimmande fiskätare. Till likheterna hörde kroppsformen, gångben omvandlade till fenor samt tanduppsättningen. Fisködlorna födde också, precis som delfinerna, levande ungar. Till skillnad från delfinerna hade fisködlorna dock bakben omvandlade till fenor samt en lodrät, inte en vågrät, stjärtfena.

I en studie från 2018 påvisade forskare ytterligare likheter med delfinerna hos ett särskilt välbevarat fisködlefossil. Dess hudyta var jämn utan fjäll och innehöll ett späcklager, precis som hos delfiner. Späck gör kroppen mer strömlinjeformad och ökar dess flytförmåga. Särskilt intressant är att späcket hos dagens valar och sälar ger värmeisolering. Därmed förhindras allt för stora värmeförluster till det omgivande vattnet, som har mycket hög värmeledningsförmåga. En låg värmeförlust är nödvändig för de vattenlevande djur som är jämnvarma ("varmblodiga") och håller en nära nog konstant kropptemperatur. Späcket tyder på att fisködlorna kanske var jämnvarma, precis som däggdjur och fåglar.
Fördelningen av pigmentkorn i fisködlefossilets hud tyder på att fisködlor, precis som delfiner, var mörka på översidan och ljusa på undersidan. Detta ger kamouflage, eftersom djuren ser vita ut underifrån mot den ljusa vattenytans bakgrund och svarta ovanifrån mot det mörka havsdjupets bakgrund. Fisködlans pigmentceller liknade däremot inte delfinernas, utan var stjärnformade som hos många reptiler. Möjligen kunde djuren skifta färg, precis som många ödlor.
På flygödlornas hud har forskarna i väl bevarade fossil identifierat grenade eller ogrenade fibrer, som skulle kunna vara en slags pälsliknande kroppsbeklädnad. Dessa fibrer har tolkats som primitiva fjädrar, men detta är kontroversiellt. I vilket fall som helst, skulle fibrerna kunna fungera som värmeisolering, precis som fåglarnas fjädrar och däggdjurens hår. Dett tyder på att flygödlorna möjligen var jämnvarma djur.

Nederst ses en modern rekonstruktion av en annan sauropod, Brontomerus. Numera anser forskare att sauropoderna var mer eller mindre "varmblodiga" (jämnvarma) och att de kunde röra sig på land. De skulle till och med kunna ha varit så alerta, att honan på bilden kunde sparka ihjäl den rovdinosaurie, som hotar hennes unge. Forskare anser numera också att sauropoderna normalt höll halsen vågrät eller endast något upphöjd i förhållande till kroppen. Utformningen av vissa sauropoders halskotor tyder på att de inte kunde höja halsen. Om de kunde höja halsen till lodrät position, kunde de förmodligen inte göra detta under någon längre tid. Då skulle hjärnans blodkärl kollapsa på grund av undertryck i dem relativt atmosfärstrycket. Alternativt skulle halsen kunna ha varit försedd med ett antal hjälphjärtan, som höjde blodtrycket till en tillräckligt hög nivå.
Fördjupning
- Om möss och elefanter: kroppsstorlekens betydelse för djurs ämnesomsättning och Hur stort kan ett djur vara och ändå kunna flyga?. Du hittar artiklarna under rubriken Varför är vissa djur stora medan andra är små.
- Trerummare eller fyrarummare? Om fördelar med olika hjärtan. Artikeln finns under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
- Djur som värmer sig med inre och yttre värmekällor: insekter, fiskar, ödlor och ormar. Läs artikeln under rubriken Att hålla sig varm när det är kallt, och kall när det är varmt – reglering av djurs kroppstemperatur.
Referenser
- M. Balter: Dinosaur metabolism neither hot nor cold, but just right (Science 344:1216-1217, 2014).
- B. L. Bostrom och D. R. Jones: Exercise warms adult leatherback turtles (Comparative Biochemistry and Physiology A 147:323-331, 2007).
- J. P. Casey, M. C. James, och A. S. Williard: Behavioral and metabolic contributions to thermoregulation in freely swimming leatherback turtles at high latitudes (Journal of Experimental Biology 217:2331-2337, 2014).
- R. A. Eagle et al. Dinosaur body temperatures determined from isotopic (13C-18O) ordering in fossil biominerals (Science 333:443-445, 2011).
- G. M. Erickson, K. C. Rogers, och S. A. Yerby: Dinosaurian growth patterns and rapid avian growth rates (Nature 412:429-433, 2001).
- P. E. Fisher et al.: Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur (Science 288:503-505, 2000).
- J. F. Gillooly , A. P. Allen, och E. L. Charno: Dinosaur fossils predict body temperatures (PLoS Biol 4[8]: e248, 2006).
- John M. Grady et al.: Evidence for mesothermy in dinosaurs (Science 344:1268-1272, 2014).
- J. Lindgren et al.: Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur (Nature 564:359-365, 2018).
- R. Motani et al.: Terrestrial origin of viviparity in Mesozoic marine reptiles Indicated by early triassic embryonic fossils (PLoS ONE 9: e88640, 2014).
- F. V. Paladino, M. P. O'Connor, och J. R. Spotila: Metabolism of leatherback turtles, gigantothermy, and thermoregulation of dinosaurs (Nature 344:858-860, 1990).
- D. J. Pierson: The physiology of dinosaurs, circulatory and respiratory function in the largest animals ever to walk the earth (Respiratory Care 54:887-911, 2009).
- G. D. Ruxton: Heat loss from giant extinct reptiles (Proceedings of the Royal Society London B 268:1921-1924, 2001).
- P. M. Sander et al.: Adaptive radiation in sauropod dinosaurs: bone histology indicates rapid evolution of giant body size through acceleration (Organisms, Diversity & Evolution 4:165-173, 2004).
- P. C. Sereno et al.: Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina (PLoS ONE 3[9]: e3303, 2008).
- M. P. Taylor et al.: Brontomerus mcintoshi, a new sauropod dinosaur from the lower Cretaceous Cedar Mountain formation, Utah, USA (Acta Palaeontologica Polonica 56:75-98, 2011).
- Z. Yang et al.: Pterosaur integumentary structures with complex feather-like branching (Nature Ecology & Evolution 3:24-30, 2019).

Har du undrat hur en fågel som står på isen en kall vinterdag undviker att förfrysa fötterna? Håller den fötterna varma eller klarar den av att ha mycket kalla fötter? Och hur är det med en fågel som står på ett hett plåttak? Hur undviker den brännskador?
Fåglar med kalla fötter
Fåglar i kalla klimat kan verkligen ha kalla ben med kalla fötter. Fottemperaturen kan understiga 5 °C, medan temperaturen i bålen är den normala cirka 40–42 °C.
Precis som vi däggdjur, så håller fåglarna en nära nog konstant och hög temperatur i kroppens kärna, det vill säga i det inre av huvudet och bålen. Men gångbenen ställer till med problem för en fågel när det är kallt. Deras nedre, obefjädrade delar är dåligt isolerade. Deras yta utgör en stor del av kroppsytan och är dessutom mycket stor i förhållande till deras volym. En stor relativ yta ger en stor värmeavgivning. En liten volym ger en liten värmeproduktion. Ett ben kan därför knappast producera tillräckligt med värme själv för att hålla en hög temperatur i en kall miljö. I stället kan det fungera som en värmeradiator, som vid kyla via blodet till omgivningen avger stora mängder av den värme som kroppskärnan producerar. Då kyler den ner hela fågeln.
Lösningen på detta problem är att hålla gångbenens temperatur väsentligt lägre än kroppskärnans. Värmeavgivningen blir nämligen mindre, om temperaturskillnaden mellan benet och omgivningen är liten. Benet värms upp av varmt arteriellt blod som flödar ner i det från bålen. Djur kan kyla benen genom att minska blodflödet till dem. När bentemperaturen minskar så minskar ämnesomsättningen och därmed benets syrgasbehov. Benet kan då klara sig med ett mindre blodflöde. Men hos många fåglar och däggdjur hittar vi värmeväxlare vid benens bas som kyler benen på ett mycket effektivare sätt.

Hur fungerar värmeväxlare i kyla?
I värmeväxlarna löper artärer med varmt syrerikt blod parallellt med vener med kallt syrefattigt blod. Artärerna löper från bålen ut till benet, venerna från benet in till bålen. En del värme överförs från det varma artärblodet till det kalla venblodet. Denna värme transporteras inte ut i benet, utan blir kvar i bålen. Bentemperaturen blir då lägre. Ju längre kärlen i värmeväxlaren är, desto mer värme kan överföras från artärer till vener. En längre värmeväxlare ger alltså en lägre bentemperatur. Avståndet mellan det arteriella och det venösa blodet i värmeväxlaren är så stort att syrgas inte kan transporteras mellan de båda blodbanorna. En värmeväxlare kan således kyla ett ben utan att minska syrgastillförseln till det. Den nödvändiga tillförseln och bortförseln av andra ämnen till och från benet påverkas inte heller. Blodkärlen i värmeväxlaren är så pass vida att blodet relativt lätt kan ta sig igenom dem. Med andra ord, så har de en relativt låg resistans. Därmed går endast en liten del av blodets tryckenergi förlorad när det passerar värmeväxlaren och det behövs inget extra hjärta i benet.
Hos vissa djur, till exempel människan, består värmeväxlaren bara av några artärer och vener, som ligger intill varandra. Vi är väsentligen tropiska djur, dåligt anpassade till att tåla kyla. Hos djur som lever i kalla klimat kan värmeväxlaren bestå av ett betydligt effektivare så kallat rete mirabile ("det underbara nätverket"). Det består av ett stort antal artärer och vener som ligger intill och blandade med varandra. I tvärsnitt ser det ut som ett nät.
Det kan vara svårt för muskler att fungera i kyla. Men det finns nästan inga muskler i de obefjädrade nedre delarna av fågelbenet. Nästan alla de muskler som påverkar benet finns i bålen eller i benets övre fjäderklädda del, där temperaturen är relativt hög. Dessa muskler manövrerar benet via långa senor som löper i skidor längs med benet. Den obefjädrade delen av benet består mest av hud, ben, brosk och bindväv. Dessa vävnader tål kyla relativt bra. Benets nedre del har också ett känselsinne. Det innehåller sinnesreceptorer som reagerar på bland annat tryck och temperatur. Det innehåller nervfibrer som förmedlar sinnesintrycken till ryggmärgen och hjärnan. Dessa nervfibrer är utskott från nervcellkroppar som finns i bålen nära ryggraden. Det är mycket märkligt att en och samma nervfiber kan fungera vid cirka 5 °C i foten och vid cirka 40 °C i bålen. Olika delar av samma nervcell är alltså anpassade till radikalt skilda temperaturer. Vi vet inte hur detta är möjligt.
Om fötterna riskerar att frostskadas eller rent av frysa till is, kan fågeln stänga av värmeväxlarna genom att leda venblodet tillbaka via ytliga vener, i stället för via värmeväxlarnas djupa vener. Då värms gångbenen upp av det varma arteriella blodet. Fågeln kan också värma upp benen genom att öka blodflödet till dem. Detta underlättas av så kallade arteriovenösa anastomoser i huden. Dessa för blod direkt från artärer till vener, utan att det passerar kapillärsystemen. Då minskar resistansen ("motståndet") mot blodflöde i huden, så att dess genomblödning ökar. Därmed värms huden snabbare och frostskador undvikes.
Värmeväxlare finns inte bara i fåglars gångben. De finns bland annat i gångben, ytteröron och svansar hos många däggdjur. Exempel är hundens gångben, valarnas fenor, hararnas öron, bäverns svans och till och med våra egna armar och ben. Fast vi klarar förstås inte av att hålla en temperatur på 5 °C i händer och fötter.

Hur fungerar värmeväxlare i hetta?
Många djur kan, när de är överhettade, stänga av värmeväxlarna och samtidigt öka blodflödet till gångbenens hud, så som beskrivits ovan. De förvandlas då till radiatorer som avger överskottsvärme från kroppen. Värmeväxlarna stängs av genom att venblodet leds tillbaka via ytliga vener, i stället för via värmeväxlarnas djupa vener. Sådana ytliga vener syns som blåa streck på våra egna armar. Dessa vener syns knappt vid köldstress, men kan svälla upp vid värmestress Hos många däggdjur i varma klimat finns dessutom väl genomblödda ytförstorade kroppsdelar som är speciellt anpassade till att fungera som värmeradiatorer i hetta. Ett exempel är elefanternas öron.
Hur kan en fågel stå på ett hett plåttak utan att få brännskador?
Här hittar jag ingen vägledning i litteraturen. Det återstå då för mig att spekulera.
Ett hett solbelyst plåttak kan troligen anta temperaturer uppemot 40–50 °C eller mera vid en lufttemperatur på 30 °C . Takets temperatur påverkas naturligtvis av solhöjden. Ju högre solen står, ju intensivare blir solstrålningen. Takets temperatur påverkas också av dess färg. Ett svart tak absorberar mer synlig solstrålning än ett vitt. Vinden har också betydelse. Vid hög vindstyrka avges mer värme från taket med så kallad konvektion. En nackdel för fågeln är att plåtens metall har en mycket hög värmeledningsförmåga.

Fågelns fötter innehåller inga muskler, utan huvudsakligen skelettben, bindväv, senor hud samt nervfibrer som mottar sinnesintryck. Generellt sett är fötternas vävnader förmodligen mindre värmekänsliga än muskler och inälvor. Fotens vävnader hos fåglar i kalla klimat har, som nämnts ovan, visat sig tåla åtminstone 5 °C . Gissningsvis klarar de också ganska höga temperaturer, men knappast mer än cirka 50 °C under längre tid. Det kan nämnas att smärttröskeln för smärtreceptorerna i människans hud ligger mellan cirka 50 och cirka 60 °C , olika i olika kroppsdelar och olika vid olika uppvärmningshastighet.
Fåglars temperatur i kroppens kärna, det vill säga i de inre delarna av huvudet och bålen är cirka 40–42 °C . Teoretiskt skulle en fågel kunna kyla fötterna genom att koppla ur värmeväxlarna i benen och kraftigt öka blodflödet till fötterna, eftersom taktemperaturen är högre än kärntemperaturen. Blodet skulle då sänka temperaturen i fötterna. Men detta skulle kunna leda till värmestress, eftersom det av taket uppvärmda blodet värmer upp kroppens kärna. Om jag vore en fågel, skulle jag hellre flyga min väg.
Är fåglarnas fotsulor isolerande som skosulor?
Det är tänkbart att vissa fåglars fötter är ganska väl isolerade mot värmetransport. Fåglar har ett tjockt hornlager på fotens undersida. Lagret består bara av dött material (hornämnen, keratiner) och leder värme ganska dåligt. Detta har visats vara av betydelse, när fåglar står på ett kallt underlag. Det bör då också vara det, när de står på ett varmt. Dessutom har många fåglar små utskott, papiller, på fotens undersida. Om foten vilar på papillerna, minskar den yta som är i kontakt med underlaget. Det finns data som tyder på att papillerna skyddar fågelfötter mot avkylning. I så fall skyddar de också mot upphettning.
Fördjupning
Om möss och elefanter: kroppsstorlekens betydelse för djurs ämnesomsättning. Du hittar artikeln under rubriken Varför är vissa djur stora medan andra är små?
Referenser
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (4th ed, Sinauer, 2018).
- P. E. Hillman och N. R. Scott: Energy budget of the chicken's foot (Journal of Thermal Biology 4:205-217, 1989).
- I. Lennerstedt: Seasonal variation in foot papillae of wood pigeon, pheasant and house sparrow (Comparative Biochemistry and Physiology 51A:511-520, 1975).
- U. Midtgård: Morphometric study of structures important for cold resistance in the arctic Iceland gull compared to herring gulls (Comparative Biochemistry and Physiology 93A:399-402, 1989).
- A. Pertovaara et al.: Influence of skin temperature on heat pain threshold in humans (Experimental Brain Research 107:497-503, 1996).

Hur ska du klä sig när det är mycket varmt, som i öknen? Ökenlevande folkgrupper, som beduiner, använder både svart och vit klädsel. Intuitivt tycker vi kanske att vit klädsel borde ge mindre värmestress. Men är det så?
Synlig strålning från solen
Solen avger både synlig strålning och värmestrålning. Vit klädsel både reflekterar och släpper igenom mer synlig solstrålning än svart. Den synliga strålning som absorberas av kläderna eller passerar igenom dem till huden omvandlas till värme. Med vita kläder borde således kroppens värmeupptag från synlig solstrålning i hög grad bero på hur stor andel av ljuset som reflekteras av kläderna i förhållande till hur stor andel som absorberas av kläder eller hud. Detta bestäms i sin tur av klädestygets egenskaper.
Om tyget är tillräckligt tjockt och tättvävt, absorberar däremot svart klädsel nästan all synlig solstrålning på sin yta, varvid ytan värms upp. Om de svarta kläderna är tätsittande kan den bildade värmen lätt nå kroppen. Om kläderna däremot är vida och sitter löst finns det mycket luft mellan dem och kroppen. Då skulle du kunna tänka dig, att den mesta värmen förs bort av vind utanför eller av luftflöde innanför kläderna, så kallad konvektion, och därför inte värmer upp kroppen.
Värmestrålning från solen
Vi kan mycket grovt säga att solstrålningsenergin under en klar himmel vid jordytan till 40 procent består av för människan synlig strålning och till 60 procent av infraröd strålning, det vill säga värmestrålning. Ur uppvärmningssynpunkt kan vi försumma andra våglängder. Klädernas färg har ingen betydelse när det gäller infraröd strålning. Nästan ingen infraröd strålning reflekteras, varken från kläder eller hud. Nästan all infraröd strålning som träffar kläder absorberas av dem. Detta gäller både vita och svarta kläder. Infraröd strålning kan eventuellt i viss utsträckning passera genom klädernas tyg till huden. Mängden strålning som passerar beror i så fall på hur pass tjockt och tättvävt tyget är, inte på dess färg.
Solstrålningens sammansättning
Luftflöde, svettning och avdunstning
Löst sittande kläder har en stor fördel, när vi svettas i värme. När vi rör oss, så rör sig också luften mellan kläderna och kroppen. Sådant konvektivt luftflöde underlättar svettens avdunstning till gasformigt vatten, alltså vattenånga, genom att föra bort den bildade ångan. När svettens vatten avdunstar på huden åtgår värme, vilket leder till att kroppen kyls. Har du en tätsittande tröja blöter svetten tröjan, i stället för att avdunsta på huden och den avkylande effekten blir sämre.
Studier av människor
Det finns några studier som tagit sig an problemet med svarta kontra vita kläder. I ett fall gjorde forskarna upprepade försök med en beduin i Negevöknen vid lufttemperaturer mellan 35 och 46 °C och låg vindstyrka, 0-4,1 meter per sekund. Han var klädd i sin traditionella klädsel, antingen en svart kåpa eller en likartad vit kåpa. Den svarta kåpan uppvärmdes som väntat mer än den vita, eftersom den absorberade mer synlig solstrålning. Båda kåporna avgav ungefär lika mycket infraröd strålning till omgivningen. Den svarta kåpan förlorade mer värme än den vita genom vindens konvektion, eftersom dess yta var varmare. Men den avkylande effekten av den svaga vinden var liten. Nettoeffekten blev att den svarta kåpan tog upp ungefär tre gånger så mycket värme som den vita. Men trots detta tog beduinen inuti kåpan upp ungefär lika mycket värme iklädd svart kåpa som iklädd vit kåpa. Och lufttemperaturen mellan kåpan och kroppen var ungefär den samma under båda kåporna. Den värmeisolerade effekten var alltså lika stor för båda kåporna. Forskarna förmodade att den höga temperaturen på den svarta kåpans insida ledde till ett större konvektivt luftflöde inuti denna kåpa än inuti den vita. Den svarta kåpan fungerade kanske som en skorsten. Detta skulle innebära att varm luft steg uppåt på grund av sin låga densitet ("täthet") och fördes ut ur kåpan upptill. Detta i sin tur ledde till att svalare luft sögs in under kåpan vid dess underkant.
I en annan studie undersökte forskarna vietnamesiska kvinnor iklädda sina traditionella kläder i solsken. Till skillnad från beduinen gick kvinnorna i normal promenadtakt. Kläderna var vita eller svarta, men i övrigt identiska. De svartklädda kvinnorna hade en lägre kroppstemperatur och en lägre temperatur mellan kläderna och kroppen än de vitklädda. De svartklädda kvinnorna var således med stor sannolikhet mindre värmestressade än de vitklädda. Även här trodde forskarna att det berodde på den ovan beskrivna skorstenseffekten.
En undersökning av cyklande danskar och danskor i artificiellt solljus gav ett annorlunda resultat. De svartklädda försökspersonerna var mer värmestressade än de vitklädda. Skillnaderna var emellertid små. Och personerna var iklädda träningsoveraller, inte sådana vida kläder som används av många folkslag under den tropiska solen.
Med denna tillgängliga information måste vi dra den förvånande slutsatsen, att rätt utformade svarta kläder sannolikt kan vara lika bra eller bättre än vita, om du vill undvika överhettning i ökensolen.
Referenser
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- B. Nielsen: Solar heat load - heat balance during exercise in clothed subjects (European Journal of Applied Physiology 60: 452-456, 1990).
- M.H. Nguyen och H. Tokura: The different effects of black and white Vietnamese Aodai folk costumes on rectal temperature and heart rate in women walking intermittently in hot and sunny environment (International Journal of Occupational Medicine and Environmental Health 13:27-38, 2000).
- A. Shkolnik et al.: Why do bedouins wear black robes in hot deserts? Nature 283:373-375, 1980).
Fördjupning
Varför är så många fåglar vita? Om fjäderdräktens färger hos fåglar. Du hittar artikeln under rubriken Förnuft, känslor och beteenden – Hur funkar hjärnan, nervsystemet, sinnen och hormoner?.

Däggdjur och fåglar är ju kända för att de håller kroppen varmare än omgivningen med hjälp av egenproducerad värme och för att de allra flesta av dem reglerar kroppens temperatur, så att den är nära nog konstant. Men det finns andra djur än däggdjur och fåglar som kan värma upp sig själva med egenproducerad värme, åtminstone i vissa kroppsdelar. Det finns också djur som kan reglera sin kroppstemperatur med hjälp av beteende, åtminstone under dygnets ljusa del.
Växelvarma och jämnvarma djur
De flesta djur reglerar inte sin kroppstemperatur. Förr kallade vi sådana djur "kallblodiga", numera växelvarma. Begreppet växelvarm inkluderar tre egenskaper. Djuren är poikiloterma, eftersom deras kroppstemperatur är lika med omgivningens temperatur och varierar med den. De är ektoterma, eftersom de saknar förmågan att värma upp sig med egenproducerad värme. De kan inte reglera sin kroppstemperatur på en bestämd nivå genom inre mekanismer. Många av dem kan dock ändra sin kroppstemperatur genom beteende, till exempel genom att vid behov uppsöka varma eller kalla miljöer.
Däggdjur och fåglar kallades förr "varmblodiga", numera jämnvarma. Nästan alla är de homeoterma, eftersom de håller sin kroppstemperatur nära nog konstant och oberoende av yttertemperaturen. De är endoterma, eftersom de kan värma upp sig med egenproducerad värme. De kan reglera sin kroppstemperatur i hela kroppen med hjälp av inre mekanismer, till exempel ökad värmeproduktion, ändrad isolering eller avdunstning av vatten. För att vara jämnvarmt behöver ett djur tre egenskaper: en hög inre värmeproduktion, en god isolering oftast genom päls, fjäderdräkt eller späck samt en effektiv temperaturreglering.


Jämnvarma djur är mindre beroende av omgivningens temperatur än växelvarma, vilket är en stor fördel. En nackdel är att de måste ha en hög energiomsättning (metabolism) för att kunna producera värme. För att omsätta energi genom cellandning måste de därför också ha en hög syrekonsumtion. Detta gör att de behöver mer föda och syre än växelvarma djur. Men den höga syrekonsumtionen ger dem också en fördel. De kan utföra intensivt muskelarbete under lång tid. Många av dem är uthålliga löpare.
Men däggdjur och fåglar är inte så unika, som du skulle kunna tro. Många andra djur kan höja sin kroppstemperatur genom att själva producera värme, åtminstone i vissa kroppsdelar eller under vissa tidsperioder. Många andra djur kan reglera sin kroppstemperatur genom sitt beteende, till exempel genom att uppsöka varma eller kalla ställen eller genom att sola vid vackert väder. Vilka djur, förutom däggdjur och fåglar, kan då höja och reglera sin kroppstemperatur?
Varma insekter
En del insekter, till exempel humlor och svärmarfjärilar, kan reglera kroppstemperaturen i mellankroppen i samband med att de värmer upp flygmusklerna vid kall väderlek. De kan producera värme genom att "huttra" med flygmusklerna. Vinghöjarmusklerna drar då ihop sig samtidigt som vingsänkarmusklerna. Eftersom de båda muskelgrupperna motverkar varandra, kommer vingarna inte att röra sig eller bara vibrera svagt. Resultatet blir muskelkontraktioner utan muskelförkortning, så kallade isometriska kontraktioner, och muskelarbetet resulterar då bara i värmeproduktion. Dessa insekter har dessutom ofta kroppen, särskilt mellankroppen, isolerad av en "päls". De reglerar temperaturen i mellankroppen. Tack vare värmeproduktionen och isoleringen kan de flyga vid låga yttertemperaturer. Då är de flesta andra insekter markbundna. Humlor och många nattfjärilar är välkända exempel på detta. Pälshåren hos insekter består dock av ett ämne som kallas kitin och inte av de keratiner (hornämnen) som utgör däggdjurens päls och fåglarnas fjädrar.

Bisamhällen kan faktiskt reglera temperaturen i hela samhället. Vi kan säga att hela samhället, åtminstone avseende temperaturreglering, utgör en "superorganism".
Märkligt är också att humledrottningar kan hålla en i det närmaste konstant kroppstemperatur när de ruvar sina ägg. Samma sak gäller honor av pytonormar och boaormar. Dessa ormhonor rullar sig runt äggen och ruvar dem på fågelvis. Humleäggen och pytonäggen har alltså förmånen att få utvecklas vid en konstant och gynnsam temperatur, precis som fågelägg.
Varma fiskar
En del fiskar kan hålla en högre temperatur än det omgivande vattnet i vissa simmuskler, i hjärnan, i ögonen eller i delar av magtarmkanalen. Det handlar bland annat om svärdfiskar, segelfiskar och tonfiskar samt om hajar tillhörande familjen Lamnidae. Några av dessa fiskar reglerar också temperaturen i de uppvärmda organen. Alla "varma" fiskar använder värmeväxlare. I sådana värmeväxlare transporteras värme från det blod som lämnar ett organ till det inkommande blodet. Därigenom tenderar värmen att stanna kvar i organet och höja dess temperatur.

En del av de "varma" fiskarna har också "värmeelement". Hos bland annat segelfiskarna har muskler som normalt styr ögats rörelser omvandlats till värmeproducerande organ. Dessa organ är tillsammans med hjärnan och ögonen omgivna av ett hölje av isolerande fett. Blodkärlen till och från de i höljet inneslutna organen är arrangerade som rören i en värmeväxlare. Därmed kommer blodcirkulationen inte att skölja bort alltför mycket av den producerade värmen. De värmeproducerande organen producerar värme genom att pumpa kalciumjoner in i det endoplasmatiska nätverket. Det endoplasmatiska nätverket utgörs av membransäckar och finns inuti alla celler. Kalciumjonerna får sedan diffundera ut igen drivna av den högre kalciumhalten inne i nätverket. Processen med inpumpning och utdiffusion upprepas hela tiden och en stor mängd kemisk energi omsätts därvid till värme. Sådana processer hos djur som bara resulterar i värmeproduktion brukar kallas "futila cykler". Men de är egentligen inte futila, alltså betydelselösa, eftersom värmeproduktionen är till fördel för djuret.

Glansfisken kan reglera kroppstemperaturen på en nära nog konstant nivå, som är högre än vattentemperaturen, dock bara cirka 3–5 °C. Det kan den göra i hela kroppskärnan, det vill säga i de delar som ligger innanför huden med underliggande vävnad. Huvudets temperatur är något högre än resten av kroppens, eftersom fisken där har ett "värmeelement" påminnande om det som i huvudtexten ovan beskrivits hos segelfiskarna. De ovan nämnda "varma" fiskarna kan inte värma upp hjärtat, men det kan glansfisken. Den värmer sig med egenproducerad värme.
Värmen produceras framför allt i de jättestora bröstmusklerna, som används när fisken simmar med hjälp av bröstfenorna. Dessa muskler är isolerade med ett tjockt lager av fettvävnad och upptar 16 procent av kroppsvikten. På röntgenbilden upptar de hela området under ryggraden mellan kraniet och den randiga bakre delen av kroppen.
Unikt för glansfisken är att den har värmeväxlare i gälapparaten. Det varma syrefattiga blodet från kroppen flyter i en mängd tunnväggiga kärl till gälarna. Det syrerika blodet från gälarna flyter i en mängd likaledes tunnväggiga kärl till alla övriga organ i kroppen. De två typerna av kärl transporterar blodet i motsatta riktningar. De löper parallellt och tätt packade med varandra. Därmed behålles värmen i kroppen genom att den transporteras över kärlväggarna från det inkommande varma blodet till det kalla blod som lämnat gälarna. Hos andra fiskar förloras all värme i gälarna och blodet som lämnar dem har samma temperatur som det omgivande vattnet.
Men värmeväxlaren borde också leda till ett problem. Syre från det inåtledande syrerika blodet borde på grund av koncentrationsskillnaden transporteras till det utåtledande syrefattiga blodet. Då skulle fiskens syreupptag avsevärt försämras. Det skulle vara intressant att veta hur glansfisken löser detta problem. Vissa fiskar med värmeväxlare i andra delar av kroppen än gälarna har speciella hemoglobiner, som släpper syre så långsamt att det inte förloras i värmeväxlaren.
Många reptiler kan genom beteende reglera sin kroppstemperatur på dagen med hjälp av yttre värmekällor, framför allt solljus. De kan på detta sätt hålla kroppens temperatur nära nog konstant under dagen. Men på natten måste de hålla samma temperatur som omgivningen. Reptilernas beteenden kan vara av många olika slag. Många ödlor rör sig fram och tillbaka mellan solen och skuggan, ut i solen när de blir kalla och in i skuggan när de blir varma. När de är i solen kan många av dem också ändra färg, så att de är mörka när de är kalla och ljusa när de är varma. En mörk ödla absorberar mer av det synliga ljuset från solen, medan en ljus ödla reflekterar mer av detta ljus. Ödlor kan också dra ihop de ytliga blodkärlen för att minska värmeavgivningen från huden och utvidga dem för att ta upp mer värme från solen. De kan pressa sig mot varma eller kalla ytor i omgivningen för att ta upp eller avge värme.
Däggdjur och fåglar
Men fåglar och däggdjur är de enda djur som kan hålla kroppens temperatur konstant och mycket hög dygnet runt utan att vara beroende av yttre värmekällor. Tre viktiga genetiska anpassningar har möjliggjort detta.
För det första, så har däggdjur och fåglar en mycket väl utvecklad förmåga att själva reglera sin kroppstemperatur via ett flertal olika inre mekanismer.
För det andra, så har fåglar och däggdjur en mycket hög energiomsättning (metabolism). Därmed kan de själva producera tillräckligt mycket värme för att upprätthålla kroppstemperaturen. Värmen är egentligen spillvärme. Den är till stor del en biprodukt av de processer genom vilka födan bryts ner för att producera kemisk energi åt kroppens celler. Den kan också bildas genom futila cykler. Ett exempel på en sådan cykel är att cellerna läcker ut extra mycket natriumjoner, som måste pumpas tillbaka igen, vilket leder till en hög värmeproduktion.
För det tredje, så har fåglar och däggdjur skaffat sig en ordentlig värmeisolering. Isoleringen utgörs förstås av päls och fjädrar. Fast egentligen är det stillastående luft inuti pälsen och fjädrarna som isolerar, inte pälsen och fjädrarna i sig. Hos en del vattenlevande fåglar och däggdjur finns denna isolerande luft kvar när de dyker. Men ofta är det ett tjockt späcklager som isolerar de vattenlevande djuren. Hos valar och sälar är pälsen reducerad eller helt borta.
Fördjupning
Reglering av kroppstemperaturen i värme och i kyla: huttrande, blodflöde, gåshud och svettning. Artikeln finns under rubriken Att hålla sig varm när det är kallt, och kall när det är varmt – reglering av djurs kroppstemperatur.
Hur sker transporter i djurens kroppar? Diffusion och massflöde. Du hittar artikeln under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
Referenser
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal physiology (3rd ed, Sinauer, 2012).
- R. M. Runcie et al.: Evidence for cranial endothermy in the opah (Lampris guttatus)(Journal of Experimental Biology 212:461-470, 2009).
- K. Schmidt-Nielsen: Animal physiology (5th ed, Cambridge University Press, 1997).
- G. J. Tattersall et al.: Seasonal reproductive endothermy in tegu lizards (Science Advances 2[1]:e1500951, 2016).
- N. C. Wegner et al.: Whole-body endothermy in a mesopelagic fish, the opah, Lampris guttatus (Science 348:786-789, 2015).
- P. Willmer, G. Stone, och I. Johnston: Environmental physiology of animals (2nd ed, Blackwell, 2005).

Alla som sett filmen "Hajen" känner till vithajen (Carcharodon carcharias). Den är en mycket stor och snabb haj som aktivt jagar stora marina däggdjur, framför allt sälar, i både varma och kalla vatten. Att den är snabb och dessutom kan jaga effektivt i kallare vatten gör den till ett av de mest framgångsrika rovdjuren i havet. En anledning till detta är att den, till skillnad från nästan alla andra fiskar, tycks vara nästan jämnvarm ("varmblodig"), precis som däggdjur och fåglar. Jämnvarma djur håller en i det närmaste konstant temperatur i kroppens kärna, men inte i de yttre delarna.
Vithajen kan, med hjälp av värme som produceras i dess egen kropp, hålla en högre temperatur än vattentemperaturen i större delen av kroppens kärna. Till detta kommer att hajen har ett stort hjärta med en muskulös kammare och ett blod med en hög halt av syrgasbindande hemoglobin, Detta är egenskaper som vi annars hittar främst hos de jämnvarma däggdjuren och fåglarna.
Vithajen har en kärntemperatur, mätt i magsäcken, som är upp till 14 °C högre än det omgivande vattnets, medan dess hud och ytligare belägna delar har ungefär samma temperatur som omgivningen. Magsäckens temperatur håller sig inom ett snävt område, även när omgivningens temperatur varierar kraftigt. Vithajen förefaller alltså reglera temperaturen i större delen av kroppskärnan. Den håller kärntemperaturen inom spannet 24-28 °C. Detta är lägre temperaturer än däggdjurens (cirka 36-38 °C) och fåglarnas (cirka 40-42 °C).

Hur kan då vithajen höja sin kroppstemperatur när nästan alla andra fiskar saknar förmågan? Svaret heter värmeväxlare. När fiskar simmar och därmed använder sina muskler bildas värme i dessa. Även i andra organ bildas spillvärme vid ämnesomsättningen. Denna värme transporteras till stor del runt i kroppen med blodet, eftersom blodet värms upp när det passerar organen. Hos nästan alla fiskar förloras all värme genast till det omgivande vattnet, när blodet når ytliga kärl, i synnerhet kärlen i gälarna. Hos vithajen sparas i stället värmen i kroppens inre. Värmeväxlare gör att värmen aldrig når de ytliga delarna av hajen och inte heller gälarna.
Vener med syrgasfattigt blod som lämnar flera av vithajens organ möter tidigt på sin väg artärer med syrgasrikt blod som går till organet. Vener och artärer löper nära varandra ett långt stycke, innan de skiljs åt. Det inåtgående venösa blodet är kallt, det utåtgående arteriella blodet är varmt. Eftersom värme alltid transporteras från ett varmt område till ett kallt, går värmen genom blodkärlsväggarna från det utåtgående syrgasfattiga blodet till det inåtgående syrgasrika blodet och därmed tillbaka till organet. En del av den värme som organet producerar kommer alltså att stanna kvar i det och höja dess temperatur.
Vithajen har värmeväxlare som försörjer muskulaturen, hjärnan, ögonen och inälvorna. Varje organ är utrustat med ett så kallat rete mirabile ("underbart nätverk") som består av ett stort antal parallella vener och artärer. Hajen har kanske till och med en förmåga att reglera vissa organs temperatur genom att stänga av värmeväxlaren när organet är för varmt och i stället använda sig av andra blodkärl som inte ligger så nära varandra som de i ete mirabile.
Förmågan att hålla en högre temperatur än omgivningen i delar av kroppen förekommer bland annat hos hajar inom familjen Lamnidae, dit vithajen hör, och många arter inom tonfiskfamiljen. De flesta av dessa fiskar har emellertid färre värmeväxlare och flera av dem kan inte hålla temperaturen på en konstant nivå.
Vilka fördelar ger då en högre inre kroppstemperatur? Genom att hålla en hög temperatur i utvalda delar av kroppen får hajen förmodligen en förhöjd aktivitet i nervsystemet och musklerna samt en snabbare nedbrytning av födan. Den förhöjda matsmältningsförmågan skulle kunna göra att hajen bättre utnyttjar energitillskottet från bytet och därför inte behöver jaga lika ofta. De uppvärmda simmusklerna kan förmodligen arbeta med större effekt. Det är kanske detta som gör vithajen till en så snabb fisk. Men den största fördelen är kanske att vithajen, tack vare sin höga kroppstemperatur, kan vara mycket aktiv i kalla havsområden.
Referenser
K.J. Goldman: Regulation of body temperature in the white shark, Carcharodon carcharias (Journal of Comparative Physiology B 167:423-429, 1997).
R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
Djurens evolution och människans ursprung – åldrande och livslängd

Vad är ett djur? Det är en grupp organismer med gemensamt ursprung. De är alla flercelliga. De härstammar alla från en organism, som troligens levde för cirka 600 miljoner år sedan. Djuren kan definieras på två sätt: genom de strukturer och molekyler, som finns hos alla djur och bara hos djur, och genom deras inbördes likheter gällande arvsmassans DNA. Det senare avser ordningsföjden för de fyra baser, som utgör bokstäverna i det genetiska alfabetet.
Molekylärbologisk definition
Begreppet djur måste alltså definieras av släktskapsförhållanden. Djur är en grupp organismer med en gemensam förfader. Släktskapen mellan organismer bedömdes tidigare enbart med hjälp av morfologiska egenskaper, det vill säga kroppens, vävnadernas och cellernas struktur, i viss mån också förekomsten av vissa molekyler. Men vi har numera tillgång till en mängd molekylärbiologiska data, framför allt bassekvenser i DNA. Sådana data har gjort det lättare att bedöma olika organismers släktskap med varandra. Med hjälp av statistiska analyser av likheter i arvsmassans DNA har forskare visat vilka organismer som är djur.De har också visat att alla djur har ett gemensamt ursprung och konstruerat ett släktträd för djuren. Men DNA-studier är ofta svårtolkade. Detsamma gäller morfologiska studier. När de båda metoderna ger olika resultat, blir det ännu svårare att bygga upp ett släktträd. Nya studier kommer troligen leda till att släktträdet förändras.
Om svampdjur och andra djur som inte förflyttar sig

Svampdjuren (Porifera) är mycket enkelt byggda med ett fåtal celltyper och inga riktiga vävnader. Traditionellt har de betraktats som den nedersta grenen på djurens släktträd, en systergrupp till alla andra djur. Men de har nu utmanats av kammaneterna. Svampdjuren sitter fast i underlaget och kan inte röra på kroppen. De filtrerar födopartiklar från det omgivande vattnet. Vattnet drivs genom kanaler i svampkroppen av tunna rörliga utskott, som kallas gissel eller flageller och finns på särskilda celler, koanocyter.
Om vi delar upp ett svampdjur i enskilda celler och placerar alla cellerna i samma kärl, så kommer cellerna att organisera sig till en ny individ. Det är tveksamt om svampdjuren har de sammanhängande cellskikt, epitel, som finns hos alla andra djur och som är nödvändiga för att bygga upp en komplicerad kropp. Svampdjuren är nästan ett mellanting mellan en koloni av enskilda celler och en flercellig organism.
Hårstjärnorna är tagghudingar, släkt med sjöstjärnor och sjöborrar. Vuxna hårstjärnor kan förflytta sig med hjälp av de fjäderliknande armarna som syns överst på bilden. Men de sitter oftast fast i underlaget med hjälp av korta utskott (cirrer). Hårstjärnornas släktingar sjöliljorna sitter som vuxna fast i underlaget hela tiden och kan bara röra på armarna. Båda grupperna filtrerar små födopartiklar ur vattnet med hjälp av armarna.
Djur är flercelliga. Det finns inga encelliga "urdjur"
Tidigare räknade forskarna många encelliga organismer, så kallade "urdjur", till djuren. Men "urdjuren" är inte alls närmare släkt med de flercelliga organismer, som kallas djur. Därför räknas numera bara flercelliga organismer till gruppen Animalia (Metazoa; djurriket). De otaliga encelliga organismerna räknas alla numera till protisterna, som emellertid inte utgör en naturlig grupp grundad på släktskap. Det finns ett stort antal protistgrupper, av vilka många står lika långt ifrån varandra som djuren står från växterna. Koanoflagellaterna är den protistgrupp som står närmast djuren.
Morfologiska egenskaper som finns hos de flesta djur
Djur kan alltså definieras molekylärbiologiskt eller morfologiskt. Vilka morfologiska egenskaper finns hos alla djur och inte hos några andra organismer? Den frågan är inte helt lätt att besvara.
Djur är flercelliga. Men det finns många andra flercelliga organismer, till exempel växter, svampar och till och med vissa bakterier. Djur är heterotrofa, vilket innebär att de får sitt energibehov tillgodosett genom att ta upp energirika kemiska föreningar, med andra ord föda. Men det finns många andra organismer som är heterotrofa, till exempel de flesta bakterier och alla svampar (det vill säga kantareller, jästsvampar och deras släktingar). De flesta djur har cellerna organiserade i vävnader och organ. Men egentliga vävnader och organ förekommer inte hos svampdjuren (till exempel tvättsvamparna). Och vävnader och organ finns också bland annat hos växterna.
De flesta djur har en matspjälkningsapparat (mag-tarmkanal) för intag och nedbrytning av föda. Men det finns undantag, till exempel svampdjur och binnikemaskar. De flesta djur har nervceller och muskelceller. Undantagen är få, men svampdjuren saknar alltid dessa celltyper. Nervceller och muskelceller finns dock bara hos djur, inte hos några andra organismer. De flesta djur har rörelseförmåga och kan förflytta sig. Det finns dock många djurgrupper som är fastsittande och saknar förmåga att förflytta sig, åtminstone i vuxet stadium. Larverna kan dock i regel förflytta sig. Även fastsittande djur har dock vanligen rörelseförmåga i någon del av kroppen med hjälp av muskelceller eller flimmerhår (cilier). Å andra sidan finns det organismer med förmåga att röra sig eller förflytta sig som inte är djur, till exempel vissa växter och bakterier.

De allra flesta djur är bilateralsymmetriska. Detta innebär att de har en framände och en bakände, en ryggsida och en buksida samt en vänstersida och en högersida. Framänden är oftast försedd med ett huvud. Dessa djur har ett enda symmetriplan ("spegelplan"). Högersidan och vänstersidan är nämligen spegelbilder av varandra. Vissa avikelser från symmetrin kan dock förekomma, ett exempel är bålens inälvor hos ryggradsdjuren.
Men alla djur är inte bilateralsymmetriska. Några enkelt uppbyggda djurstammar nära släktträdets bas är det inte. Nässeldjuren (stammen Cnidaria; bland andra maneter och koraller) är radialsymmetriska, vilket innebär att de har flera symmetriplan, precis som ett hjul. Ett hjul kan ju skäras i två delar rakt genom navet på flera olika sätt, som alla resulterar i två bitar vilka är spegelbilder av varandra. Kammaneterna (stammen Ctenophora) är biradialsymmetriska, vilket innebär att de har två symmetriplan. De enklast byggda djurstammarna, plakozoer (Placozoa) och svampdjur (Porifera) saknar helt symmetriplan.
Morfologiska egenskaper som finns hos alla djur och bara hos djur
Hittills har vi kommit fram till många morfologiska egenskaper, som finns hos de flesta eller nästan alla djur. Men vi har inte kommit fram till några morfologiska egenskaper, som karakteriserar alla djur och inte finns hos några andra organismer. Det finns dock flera sådana så kallade synapomorfier. Alla djur, men inga andra organismer, har i sin så kallade extracellulära matrix, mellan cellerna, trådformade proteiner som kallas kollagener. Kollagenerna bidrar till att ge vävnaderna mekanisk hållfasthet. Alla djur har vissa typer av cellkontakter, som inte finns hos andra organismer. Sådana kontakter kan binda ihop vävnadscellerna eller hindra transport genom cellmellanrummen i ett cellskikt. Alla djur har med stor sannolikhet ett mer eller mindre klotformat embryonalt stadium som kallas blastula. Blastulan kan dock se olika ut hos olika djurgrupper. Det finns också grupper vars embryonala utveckling inte har studerats. Blastulan bildas efter det befruktade äggets första celldelningar, klyvningsdelningarna. Blastulastadiet är unikt för djuren. Det finns också andra karaktärsdrag i könscellernas bildning och i embryots utveckling, som finns hos alla djur och saknas hos andra organismer.

Hox- och ParaHox-gener. Släktträdets grenar
Bilateralsymmetriska djur har en eller flera uppsättningar av så kallade Hox-gener samt dessutom med dessa besläktade ParaHox-gener. I en uppsättning Hox-gener sitter oftast flera Hox-gener med olika funktioner i rad efter varandra längs med DNA-molekylen i en kromosom. Under embryonalutvecklingen styr dessa gener utvecklingen av de olika kroppsdelarna utmed kroppens längdaxel, mellan framänden och bakänden. Generna sitter i samma ordningsföljd som kroppdelarna de påverkar.
Hox- och ParaHox-gener finns bara hos djur. De finns inte hos djurens närmaste släktingar, koanoflagellaterna. Hos djurstammar nära basen av djurens släktträd har forskarna funnit färre Hox- och ParaHox-gener än hos mera komplicerat byggda grupper. Hos några enkelt byggda bilateralsymmetriska maskar (gruppen Xenacoelomorpha) och hos nässeldjuren har forskarna bara hittat ett fåtal Hox- och ParaHox-gener. Hos plakozoerna har de bara hittat en sådan gen, troligen en ParaHox-gen. Tidigare trodde forskarna att svampdjuren saknade Hox- och ParaHox-gener, men de har nu hittat en ParaHox-gen hos ett svampdjur. Efter studier av DNA-sekvenser som omger Hox-gener har forskarna dessutom dragit slutsatsen att förfäder till dagens plakozoer och svampdjur förmodligen hade Hox-gener. Dessa nu försvunna gener har de kallat "spökgener".
Men det finns en djurgrupp som, så vitt vi nu vet, saknar Hox- och ParaHox-gener, nämligen kammaneterna. Detta och andra molekylärbiologiska data har lett till en intensiv kontrovers om deras ställning på djurens släktträd. Flera, men inte alla, de molekylärbiologiska studierna av djurens släktträd tyder på att de relativt komplicerat uppbyggda kammaneterna är den nedersta grenen på djurens släktträd, en systergrupp till alla andra djur. I så fall har nervceller och muskelceller under djurens evolution uppkommit vid två tillfällen, oberoende av varandra. Morfologin tyder däremot på att svampdjuren, som saknar nervceller och muskelceller, står närmast djurens förfader. Då utgör de den nedersta grenen och plakozoerna nästa gren. I så fall uppträdde kammaneterna betydligt senare under djurens evolution som en systergrupp till nässeldjuren.
Det finns andra problem med djurrikets släktträd. Det finns många gener som entydigt karakteriserar dem. De icke bilateralsymmetriska stammarna svampdjur (Porifera), plakozoer (Placozoa), kammaneter (Ctenophora) och nässeldjur (Cnidaria)) är djur. Dessa fyra djurgrupper betraktas vanligen som ursprungligare än de bilateralsymmetriska djuren. Men det finns andra möjligheter, inte bara avseende kammaneterna. De bilateralsymmetriska djuren (Bilateria) utgör återstoden av djurvärlden och upptar därför huvuddelen av släktträdets grenverk. De indelas i ett stort antal stammar.
Ett stort problem vid konstruktionen av djurens släktträd är frågan om en enkelt uppbyggd djurgrupp härstammar från förfäder med enkel uppbyggnad eller om förfäderna haft en mera komplicerad uppbyggnad. I det senare fallet kan djurgruppen under evolutionens gång ha förlorat bland annat organ, celltyper och gener. Då är den bara skenbart en ursprunglig djurgrupp. Det finns flera belagda fall av skenbart ursprungliga djur. En del menar att svampdjuren, plakozoerna och de xenacoelomorfa maskarna är sådana skenbart ursprungliga djur. I så fall skulle förfadern till alla djur kunna ha varit en förhållandevis komplicerat uppbyggd organism med många Hox- och ParaHox-gener.
Det finns ytterligare några enkelt byggda organismgrupper, som troligen är djur. På svenska kallas de, tillsammans med plakozoerna, ofta felaktigt enskiktsdjur. De är mycket svårtolkade. Flera av grupperna är parasiter. Många anser att dessa grupper härstammar från mera avancerade djur och att deras enkla byggnad orsakats av den reduktion av organ och vävnader, som är vanlig hos parasitiska djur. Den icke parasitiska gruppen Monoblastozoa har iakttagits en gång år 1892 i kulturer från saltavlagringar i Argentina (!). Forskarna anser numera att den bara existerade i upptäckarens fantasi.
Koanoflagellaterna, djurens närmaste släktingar
De encelliga koanoflagellaterna är en utomordentligt intressant grupp bland protisterna. Likheten mellan koanoflagellaterna och svampdjurens så kallade koanocyter, se bilderna nedan, gjorde att forskarna mycket tidigt misstänkte att de är djurens närmaste nu levande släktingar. Detta har nu bekräftats genom molekylärbiologiska DNA-studier. Koanoflagellaterna är dock inte djuren förfäder. Särskilt intressant är att koanoflagellaterna kan bilda kolonier, ett förstadium till flercelliga organismer. Intressant är också att de är utrustade med gener för proteiner, som hos djur förmedlar kontakt mellan celler, och proteiner, som ingår i djurens utanför cellerna belägna så kallade extracellulärsubstans. Dessa gener har bara hittats hos de encelliga koanoflagellaterna och de alltid flercelliga djuren. Enligt en teori utvecklades de första djuren ur kolonier av koanoflagellatliknade encelliga organismer.

Referenser
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).
- M. Eitel et al.: Global diversity of the Placozoa (PLoS ONE 8:e57131, 2013).
- D. E.K. Ferrier: The origin of the Hox/ParaHox genes, the Ghost Locus hypothesis and the complexity of the first animal (Briefings in Functional Genomics 15:333-341, 2016).
- D. E. K. Ferrier och P. W. H. Holland: Ancient origin of the Hox gene cluster (Nature Reviews Genetics 2:33-38, 2001).
- A. Hejnol et al.: Assessing the root of bilaterian animals with scalable phylogenomic methods (Proceedings of the Royal Society B 276:4261-4270, 2009).
- L. Margulis och M. J. Chapman: Kingdoms and domains, an illustrated guide to the phyla of life on earth (4th ed, Academic Press, 1998).
- E. Moreno, J. Permanyer, och P. Martinez: The origin of patterning systems in Bilateria – insights from the Hox and ParaHox genes in Acoelomorpha (Genomics Proteomics Bioinformatics 9:65-76, 2011).
- L. L. Moroz et al.: The ctenophore genome and the evolutionary origins of neural systems (Nature 510:109-114, 2014).
- C. Nielsen: Animal evolution, interrelationships of the living phyla (3rd ed, Oxford University Press, 2012).
- T. Nosenko et al.: Deep metazoan phylogeny, when different genes tell different stories (Molecular Phylogenetics and Evolution 67:223-233, 2013).
- O. M. Ramos, D. Barker, och D. E. K. Ferrier: Ghost loci imply Hox and ParaHox existence in the last common ancestor of animals (Current Biology 22:1951-1956, 2012).
- B. Schierwater, D. de Jong och R. DeSalle: Placozoa and the evolution of Metazoa and intrasomatic cell differentiation (International Journal of Biochemistry & Cell Biology 41:370-379, 2009).
- P. Simion et al.: A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals (Current Biology 27:1-10, 2017).
- M. Srivastava et al.: The Trichoplax genome and the nature of placozoans (Nature 454:955-960, 2008).
- J. Taylor Cannon et al.: Xenacoelomorpha is the sister group to Nephrozoa (Nature 530:89-93, 2016).
- Maximilian J. Telford, L. L. Moroz, och K. M. Halanych: A sisterly dispute (Nature 529:286-287, 2016).

Är människan ett djur? Innan utvecklingslärans genombrott intog människan en särställning bland alla de levande organismerna. Numera har vi förstått att människan är ett djur, låt vara med vissa exceptionellt välutvecklade egenskaper. Till dessa hör medvetande, kognitiv förmåga ("intelligens"), språk och kultur. Dessa egenskaper finns emellertid också hos andra djur, om än mindre väl utvecklade eller bara som ansatser.
Människan är ett djur. Bevisen för detta är överväldigande. Ser du på hur celler och organ fungerar hos människor så är likheterna med andra varelser otaliga och slående. I all synnerhet gäller detta de organismer vi kallar djur. Inte ens om du avvisar utvecklingsläran, kan du komma ifrån att människan måste vara ett djur.
Skillnaderna mellan människan och andra djur är mindre än vi skulle kunna tro
En mängd studier av andra djur har dessutom visat att medvetande, "intelligens", språk och kultur inte är unika för människan utan sannolikt har utvecklats gradvis under evolutionens gång. De hos människan exceptionellt välutvecklade kognitiva förmågorna ("intelligensen") är således inte egenskaper som plötsligt uppkommit hos vår art. De är utvecklingar och kombinationer av mekanismer, som finns även hos andra däggdjur, om än mindre väl utvecklade. Känslomässigt, emotionellt, skiljer vi oss med största sannolikhet mycket lite från andra däggdjur. En viktig insikt är således att det finns biologiska grundvalar för människans natur och för hennes beteende, både för hennes goda och hennes onda handlingar. Människor handlar rationellt, förnuftsmässigt, i betydligt mindre utsträckning än vad vi själva tror. Ofta vägleds vi av icke medvetna känslomässiga beslut. En rad psykologiska och neurobiologiska studier ger stöd åt ovanstående bild av människans villkor.
Biologer är inte biologister
Men det finns fortfarande de som tror att evolutionsforskningens och neurofysiologins rön inte kan tillämpas på människan, att hon från födseln mentalt är ett i stort sett oskrivet blad, som nästan helt formas av sin miljö, eller att hon alltid kan agera förnuftsmässigt. Trots att vi numera skaffat oss en allt bättre förståelse av hjärnan, tror vissa att våra mentala funktioner, som vilja och medvetande, inte är biologiskt grundade. De tror med andra ord att det finns en "själ", som existerar oberoende av kroppen och följer sina egna lagar.
De som har sådana åsikter beskyller ofta biologer för så kallad "biologism". Detta begrepp används på olika sätt. Men det tolkas ofta som att människan till sin natur är helt eller till allra största delen styrd av ärftliga faktorer och att hennes "biologiska" egenskaper ska vägleda oss i våra etiska ställningstaganden. För biologer är detta rent nonsens. I mer än hundra år har biologer vetat att människor, liksom andra djur, formas av ett komplicerat samspel mellan arv och miljö.

Biologi och etik
Människan är ett djur. Men hennes värdighet förminskas inte av detta. Och det leder inte heller till några som helst konsekvenser för hennes etiska värderingar. Våra etiska värderingar är ju våra uppfattningar om vad som är rätt och fel i vårt handlande gentemot andra människor och gentemot andra djurarter. Etiska värderingar kan inte grundas på biologi. Biologin kan däremot i mycket hög grad hjälpa oss att förklara människans natur, både hennes onda sidor och hennes goda. Denna kunskap hjälper oss, när vi försöker förbättra människors levnadsvillkor och minska lidande. Men biologin kan inte tala om för oss vad som är moraliskt rätt och vad som är moraliskt fel.
Biologi och religion
Hur förhåller sig då biologin till gudstro? Själv ser jag flera hinder som omöjliggör tron på en allsmäktig och allgod skapare. Men jag respekterar dem som har en sådan tro. Och till de hinder jag ser hör inte det faktum att människan är ett djur som undergått en biologisk utveckling. Tvärtom syns det mig förmätet att hävda att människan har en särställning grundad på ett gudomligt ingripande.
Bevisen för evolutionsläran är överväldigande
När det gäller evolutionsläran så är stöden för denna teori överväldigande. Vi har ett stort antal serier av fossila djur som klart belyser hur många djurgrupper och anatomiska strukturer har uppkommit. Forskare har kunnat observera och efterbilda de mekanismer genom vilka arter uppkommer. En enorm mängd molekylärbiologiska data om organismernas gener stödjer evolutionsläran. De olika organismgruppernas släktskap och utbredning på land kan förklaras med hjälp av evolutionsläran tillsammans med kontinenternas rörelser under miljontals år. Flera olika mätmetoder ger klara bevis för att den evolutionära tidskalan rör sig om miljarder år.

Kreationism och intelligent design är inte trovärdiga
Från företrädare för "kreationism" och "intelligent design", som inte är vetenskaper, har framförts att vissa biologiska strukturer är så komplexa att de inte kan ha uppkommit genom evolution via ett antal mellanstadier ("irreducible complexity"). De exempel de ger har vederlagts. Antingen finns det mellanstadier eller så är det lätt att konstruera hypotetiska mellanstadier. Exempel är ögat, bakterieflagellen, komplementsystemet och bombarderbaggen.
Evolutionsläran kan både förklara och förutsäga observationer
Evolutionsläran är i viss mån en historisk vetenskap. Det innebär att vi inte har några direkta bevis för att den storskaliga biologiska utvecklingen ägt rum. Men de fakta vi har är mycket övertygande. Molekylärbiologiska studier av DNA har gett ett stort antal resultat som bara kunnat förklaras med hjälp av evolutionsteori. Vi har också kunnat förutsäga en flertal evolutionära utvecklingsförlopp: kända exempel är övergången från reptiler till fåglar, från vattenlevande fiskar till landlevande fyrfotadjur och från reptilliknande djur med ett hörselben till däggdjur med tre hörselben. En teori som kan förutsäga observationer står mycket stark. De vetenskaper som behandlar den mänskliga kulturens utveckling är också historiska. Vi kan inte direkt bevisa att Julius Caesar och kejsar Augustus har existerat, men med utomordentligt stor sannolikhet gjorde de det. Människan och schimpansen har med mycket stor sannolikhet en gemensam förfader som levde för flera miljoner år sedan. Hur skulle detta kunna göra människan mindre mänsklig?

Fördjupning
- Varifrån kommer vi? Om lansettfiskar, ryggradsdjur och människans ursprung. Du hittar artikeln under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Vart är vi på väg? Utvecklas människan fortfarande idag? Kan vi förutsäga hennes framtida evolution? Läs artikeln under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Återspeglar fosterutvecklingen evolutionen? Om embryots utveckling hos människan och andra djur. Artikeln finns under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Vad är djur? Vad skiljer dem från andra organismer? Hur uppkom djuren? Under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd kan du läsa artikeln.
Referenser
- Evolutionsteori.se nedladdat 15 maj 2017).
- B. Forsman: Arvet från Darwin - religion, människa, moral (Fri tanke förlag, 2009).
- Alison Gopnik: How animals think (The Atlantic, maj 2016).
- D.-E. Nilsson och S. Pelger: A pessimistic estimate of the time required for an eye to evolve (Proceedings of the Royal Society of London B 256:53-58, 1994).
- E. V. Nilsson: Vetenskapspedagogen (nedladdat 15 maj 2017).
- Panda's Thumb (hämtad15 maj 2017).
- S. Pinker: Ett oskrivet blad och andra myter om människans natur (Natur och kultur, 2006).
- TalkOrigins Archive (hämtad 15 maj 2017).
- G. Vallortigara et al.: Animal cognition (Wiley Online Library , Wiley Interdisciplinary Reviews: Cognitive Science 1:882-893, 2010).
- F. de Waal: To each animal its own cognition (The Scientist, maj 2016).
- Lars Werdelin: Människans evolution (Fauna och Flora 111:2-10, 2016).
- Engelska länkar som motbevisar begreppet "irreducible complexity", framfört som argument för "intelligent design": komplementsystemet och bombarderbaggen.

Jag fick en fråga som lydde: "Jag samtalade med en lärare som undervisar i religion och historia. Han sade att Första Moseboken har samma följd av hur allting skapades som evolutionsteorin. Första mosebok skrevs ju för 2 500–3 000 år sedan. Kunde de vara så tidigt ute? Anade människorna redan då hur det hängde ihop?"
Som biolog ger jag gärna min syn på detta. Jag har läst igenom de båda skapelseberättelserna i Första Mosebokens kapitel 1 och 2. De avviker på ett mycket djupgående sätt från den väl belagda evolutionshistorien och från naturvetenskapligt tänkande. De är helt och hållet mytologiska förklaringar av världens uppkomst, dock mycket poetiska sådana med högt litterärt värde.

Den moderna människan har förmodligen i alla tider och i alla kulturer känt ett behov att förklara världens uppkomst och människans villkor. De flesta kulturer har därför skapat religiösa myter som fyllt detta behov. I vår tid har naturvetenskapen till stora delar och på ett mycket övertygande sätt förklarat universums utveckling och de levande organismernas evolution.
Den ordning i vilken de levande varelserna skapades i skapelseberättelserna stämmer inte alls överens med det evolutionära stamträd som numera är synnerligen väl belagt.
Bibelns första skapelseberättelse
I den första skapelseberättelsen beskrivs hur de levande varelserna skapades av Gud under loppet av sex dagar. Gud skapade först landväxterna. Därefter skapade han samtidigt alla vattenlevande varelser och alla fåglar. Sedan skapades de landlevande djuren. Därefter skapade Gud mannen och kvinnan.

Vi bortser från de encelliga organismerna, som ju inte var kända när bibeln skrevs. De flercelliga organismernas evolution skedde i en helt annan ordning än den som beskrivs i den första skapelseberättelsen: först utvecklades marina växter och djur i haven, sedan landlevande växter och djur. Fåglarna uppträdde på ett sent stadium i de landlevande djurens utveckling.
Bibelns andra skapelseberättelse
Den andra och kanske äldsta skapelseberättelsen avviker i ännu högre grad från den evolutionära historien. I den skapade Gud allra först människan (hebreiska "adám") "av jord från marken och blåste in liv genom hennes näsborrar, så att hon blev en levande varelse". Den första människan var en man, senare i texten benämnd Adam. Sedan skapade Gud "en trädgård, österut i Eden" åt Adam och därefter "alla markens djur och alla himlens fåglar". Slutligen skapades kvinnan ur ett av Adams revben.

I ett ovanligt välbevarat fossil av Dickinsonia (den blå "plattan" till vänster om "bägaren" i figuren ovan) fann forskare år 2018 höga halter av kolesterol, en så kallad sterol som finns i cellernas membraner hos djur, men inte i andra organismers membraner. I bergarten utanför fossilet dominerade en annan sterol, som är typisk för grönalger. Detta tyder på att Dickinsonia var ett djur. Efter den kambriska explosionen dröjde det många miljoner år innan växter och djur uppträdde på land. "Jorden var öde och tom ..." (1 Mos 1:1), men haven sjöd av liv.
Skapelseberättelserna återspeglar den tid då de skrevs
Skapelseberättelserna synes mig framför allt återspegla en indelning av världen i fast land, flytande vatten samt luft eller himmel. Denna indelning är uppenbar och säkert urgammal. Några hundra år efter det att Gamla testamentet skrivits började grekiska naturfilosofer, under 500-talet före vår tideräkning, formulera de första icke religiöst grundade teorierna om naturens sammansättning. Detta var naturvetenskapens gryning. Empedokles hävdade att världen bestod av fyra element: jord, vatten, luft och eld. Kinesiska filosofer framlade ungefär samtidigt en liknande teori med fem element: trä, eld, jord, metall och vatten.
Skapelseberättelserna och evolutionsläran
Ur biologisk synpunkt har skapelseberättelserna dessutom andra grundläggande problem.
Ett uppenbart problem är naturligtvis Gamla testamentets tidsskala. Den geologiska tidsskalan är säkerställd genom flera olika mätmetoder. Planeten Jorden bildades för cirka 4,6 miljarder år sedan. De första flercelliga organismerna uppträdde för mer än 600 miljoner år sedan.

Ett annat problem är att skapelseberättelserna är teleologiska, vilket innebär att skapelsen är styrd mot ett mål. Gud skapade världen med ett syfte, en förutbestämd plan. Evolutionen, däremot, är inte riktad mot ett bestämt mål. Förenklat uttryckt så sker evolutionen huvudsakligen genom att vissa slumpmässiga förändringar, mutationer, i individers arvsmassa etableras genom naturligt urval. Naturligt urval sker genom att bärarna av gynnsamma mutationer är bättre anpassade till sin miljö och därför producerar en mer talrik avkomma. Det går inte att förutsäga evolutionens riktning. Vid slutet av kritaperioden dog dinosaurierna, utom fåglarna, ut. Orsaken var ett väldigt meteoritnedslag, ett enormt vulkanutbrott eller båda dessa händelser tillsammans. Om detta inte skett, så hade däggdjuren kanske förblivit en liten djurgrupp och dinosaurierna fortfarande varit de dominerande stora landdjuren.
En viktig konsekvens av evolutionens mekanismer är att de arter som uppkommer inte är "perfekt konstruerade". De är helt enkelt de arter som har överlevt och de kommer dessutom senare att ersättas av andra arter. Denna syn återfinns inte i skapelseberättelserna. I stället synes det mig att Gud drabbas av hybris inför sitt fullbordade verk: "Gud såg att allt som han hade gjort var mycket gott. Det blev kväll och det blev morgon. Det var den sjätte dagen". Den allsmäktige var märkligt blind för all denna världens ondska och lidande.
I skapelseberättelserna utmålas människan som jordens härskare. När Gud skapat mannen och kvinnan sade han till dem: "Var fruktsamma och föröka er, uppfyll jorden och lägg den under er. Härska över havets fiskar och himlens fåglar och över alla djur som myllrar på jorden." I dessa miljöförstöringens tider har vi blivit smärtsamt medvetna om att vi inte alls härskar över jorden. Vi är bara en del av den stora väven.
Fördjupning
- Betydelserna av ordet varför: om funktion, evolution och giraffens hals. Du hittar artikeln längre ner under den här rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Rudiment: evolutionära rester i människans kropp som förlorat sin funktion. Artikeln finns under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?.
- Vart är vi på väg? Utvecklas människan fortfarande idag? Kan vi förutsäga hennes framtida evolution? Du hittar artikeln längre ner under den här rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Återspeglar fosterutvecklingen evolutionen? Om embryots utveckling hos människan och andra djur. Du hittar artikeln längre ner under den här rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Är människan ett djur? Om biologi, evolution och etik.Du hittar artikeln längre upp under den här rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
Att läsa
- I. Bobrovskiy et al.: Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals (Science 361:1246-1249, 2018).
- S. A. F. Darroch et al.: Biotic replacement and mass extinction of the Ediacara biota (Proceedings of the Royal Society B 282:20151003, 2015).
- M. Z. Donahue: Did mystery worms cause world's first mass extinction? (Smithsonian Insider, uppdaterad 30 september 2015).
- Evolution (Nationellt resurscentrum för biologi och bioteknik, hämtat 13 oktober 2017).
- Evolution (Naturhistoriska riksmuseet, uppdaterat 11 februari 2016).
- Första Moseboken 1-2 (Bibel 2000).
- Första Moseboken 1-2 (1917 års översättning).
- History of life through time (University of California Museum of Paleontology, hämtad 13 oktober 2017).
- T. G. Hultman: Svensk Akademiens språklära (Norstedts, 2003).
- L. Werdelin: Människans evolution (Fauna och Flora 111:2-10, 2016).
- Understanding Evolution (University of California Museum of Paleontology, hämtad 13 oktober 2017).
- What does it mean to be human? (Smithsonian Institution, uppdaterad 13 oktober 2017).

Kanske såg det första bilateralsymmetriska (tvåsidiga) djuret, en mycket avlägsen förfader till oss, ut ungefär som masken på bilden ovan. Men vi har inga som helst fossil som visar detta enormt viktiga evolutionära framsteg. Kanske hände det så långt tillbaka i tiden som för 800 till 700 miljoner år sedan.
Masken på bilden ovan tillhör stammen Xenacoelomorpha. De flesta av dessa maskar räknades förr till stammen plattmaskar (Platyhelminthes). Numera räknas de alla vanligen som en systergrupp till alla andra bilateralsymmetriska djur. De placeras därmed vid basen av släktträdet för gruppen Bilateria, de bilateralsymmmetriska djuren. Molekylärbiologiska studier av DNA stödjer starkt denna placering. Det finns dock motsägande studier. Xenacoelomorpha omfattar tre grupper: ädelstensmaskar (Acoela), tvåstensmaskar (Nemertodermatida) och paradoxmaskar (Xenoturbellida).
Fördelar med bilateralsymmetri (tvåsidighet)
Bilateralsymmetriska eller tvåsidiga djur har en huvudände och en bakände, en ryggsida och en buksida samt en högersida och en vänstersida. Genom ett sådant djur kan du bara skära ett enda snitt, som delar djuret i två hälfter, vilka är spegelbilder av varandra. Snittet är lodrät från rygg till buk och från huvud till bakände. Det delar djuret i en högerhalva och en vänsterhalva. Bilateralsymmetri gör det lättare för djur att förflytta sig på ett underlag och att simma. Om de dessutom har mun, hjärna och sinnesorgan organiserade i ett huvud (så kallad cefalisering), blir det ännu lättare för dem att röra sig och också att skaffa sig föda.

Bilateralsymmetrin var ett avgörande evolutionärt framsteg. De flesta djur är bilateralsymmetriska. De viktigaste undantagen är svampdjur, plakozoer, nässeldjur (bland annat maneter och koralldjur) samt kammaneter.
De enklaste bilateralsymmetriska djurens byggnad
Xenacoelomorpha är de enklast uppbyggda bilateralsymmetriska djuren. De har de tre cellskikt, groddblad, som bygger upp kroppen hos Bilateria: ektoderm, mesoderm och endoderm. Ektodermet bildar hos bilaterier bland annat hud och nervsystem, endodermet mag-tarmkanalen och mesodermet flera vävnader och organ mellan de två andra skikten. Mesodermet saknas hos de ursprungligare djurstammar som står närmare basen för alla djurs släktträd: svampdjur, plakozoer, nässeldjur och kammaneter. Många menar dock att kammaneterna har mesoderm. Alla bilateralsymmetriska djur har mesoderm.

Xenacoelomorpha har förvisso de tre groddbladen, men organen är ofta inte välutvecklade och ofta ganska få. De har en framände, men den kan knappast alltid kallas huvud, se dock Symsagittifera i bilden ovan. I Proporus framände, till höger på bilden överst på sidan, ses svalget vars mynning fungerar som både mun och anus, men denna mask har inget välutvecklat huvud. I övrigt saknar djuret hålrum i kroppen. I stället för mag-tarmkanal har Proporus en central massa av sammansmälta endodermala celler, som tar hand om födan. Vissa andra maskar inom gruppen Xenacoelomorpha har en enkelt uppbyggd, blint slutande tarm.
Utsöndringsorgan, motsvarande våra njurar, saknas hos Xenacoelomorpha, men finns hos alla andra Bilateria, i sin enklaste form som så kallade protonefridier. Xenacoelomorpha har en ektodermal hud. Deras nervsystem består av nätverk av nervceller i kroppen särskilt i anslutning till huden. En del av dem har också framtill en klunga av nervceller, som utgör en enkel hjärna. Gruppen har även mesodermala muskelceller bland annat under huden och runt svalget. Musklerna kan ändra kroppens form. De rör sig dock huvudsakligen med hjälp av flimmerhår (cilier) på bukens hudceller. Könsorganens har en mycket varierande uppbyggnad.
Gener som styr embryots utveckling
I en studie från 2018 visade forskarna att genomet hos Xenacoelomorpha innehåller alla de elva genklasser av så kallade homeotiska gener, som styr embryonalutvecklingen och därmed kroppens byggnadsplan hos bilaterier. Forskarna antog att dessa genklasser fanns redan hos bilateriernas förfader. I så fall skulle den verktygslåda, som möjliggör utveckling av nya djurgrupper med radikalt annorlunda byggnadsplaner funnits långt innan den så kallade kambriska revolutionen för cirka 500 miljoner år sedan. Då uppträdde i ett geologiskt perspektiv mycket plötsligt de flesta nutida djurstammar på den fossila arenan. Detta stöder tanken att dessa djurstammar hade en lång evolutionär historia innan den kambriska revolutionen. Men forskarna har hittat mycket få eller inga fossil från prekambrisk tid, delvis kanske beroende på att djuren ännu inte utvecklat skal och skelett, hårda strukturer som lätt bevaras som fossil.
Avvikande tolkningar
Enligt en avvikande åsikt är Xenacoelomorpha inte ursprunglig. I så fall har den förlorat en rad anatomiska strukturer och härstammar från djur med mera komplicerad kroppsbyggnad. Släktskapen mellan de tre undergrupperna inom Xenacoelomorpha har också ifrågasatts.
Fördjupning
Vad är djur? Vad skiljer dem från andra organismer? Hur uppkom djuren? Du hittar artikeln längre upp under den här rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
Radiär och bilateral symmetri hos djur. Tvåsidiga djur. Varför har vi höger och vänster kroppshalva? Artikeln finns under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?.
Referenser
- J. G. Achatz et al.: The Acoela: on their kind and kinships, especially with nemertodermatids and xenoturbellids (Bilateria incertae sedis) (Organisms, Diversity & Evolution 3:267-286, 2013).
- Jaume Baguñá et al.: Back in time: a new systematic proposal for the Bilateria (Philosophical Transactions of the Royal Society B 363:1481-1491, 2008).
- M. Brauchle et al.: Xenacoelomorpha survey reveals that all 11 animal homeobox gene classes were present in the first bilaterians (Genome Biology and Evolution 10:2205-2217, 2018).
- J. T. Cannon et al.: Xenacoelomorpha is the sister group to Nephrozoa (Nature 530:89-93, 2016).
- Brenda Gavilán, E. Perea-Atienza, och P. Marínez: Xenacoelomorpha: a case of independent nervous system centralization? (Philosophical Transactions of the Royal Society B 371:20150039, 2016).
- U. Jondelius et al.: How the worm got its pharynx: phylogeny, classification and Bayesian assessment of character evolution in Acoela (Systematic Biology 60:845-871, 2011).
- U. Jondelius et al.: The Nemertodermatida are basal bilaterians and not members of the Platyhelminthes (Zoologica Scripta 31:201-215, 2002).
- H. Nakano et al.: A new species of Xenoturbella from the western Pacific Ocean and the evolution of Xenoturbella (BMC Evolutionary Biology 17:245, 2017).
- C. Nielsen: Animal evolution, interrelationships of the living phyla (3rd ed, Oxford University Press, 2012).
- H. Philippe et al.: Acoelomorph flatworms are deuterostomes related to Xenoturbella (Nature 470:255-258, 2011).

Vissa strukturer som funnits hos våra evolutionära förfäder kan anläggas i embryot under fostertiden, för att sedan försvinna. Valfoster anlägger bakbensknoppar som sedan försvinner. Fungerande gälspringor och gälbågar fanns hos våra förfäder bland fiskarna. Men gälfickor och gälbågar bildas även hos foster av människor och andra fyrfotadjur. Gälfickorna förbinds dock inte med svalget. Gälspringor bildas således inte. Gälfickorna och gälbågarna finns kvar under en period, men försvinner sedan.
Flera strukturer i huvudregionen härstammar dock från gälbågarna, hos däggdjuren bland annat mellanörat med dess ben (hammaren, städet och stigbygeln), bisköldkörtlarna, brässen (thymus), tungbenet (hyoidbenet), delar av struphuvudet samt delar av ansiktsmuskulaturen.
Det är mycket viktigt att framhålla att embryots utveckling inte strikt återspeglar ett djurs evolution. Alla organ och strukturer hos de evolutionära förfäderna återfinns således inte i ett embryo. Men vi kan ändå belysa vissa strukturers evolution genom att studera deras utveckling i embryot. Detta forskningsområde kallas numera "evo-devo".
Placentadäggdjur, pungdjur och kloakdjur
Placentadäggdjurens, inklusive människans, tidiga embryonalutveckling är ett intressant exempel på att vissa stadier återspeglar evolutionen, medan andra inte gör det. Placentadäggdjur (moderkaksdäggdjur, äkta däggdjur) och pungdjur föder levande ungar, som får näring av modern via en moderkaka (placenta). Men båda grupperna härstammar från äggläggande djur. Kloakdjuren (näbbdjuret och myrpiggsvinen) är däggdjur som fortfarande lägger ägg. Låt oss jämföra de tidiga embryonas utveckling hos placentadäggdjuren med utvecklingen hos de äggläggande släktingarna kräldjur, inklusive fåglar, och kloakdjur.
Embryots tidiga utveckling hos djur
Hos alla djur börjar den befruktade äggcellens utveckling med celldelningar som kallas klyvningsdelningar. Hos de flesta djurgrupper delas då hela äggcellen upp i flera celler, så att en klump, morula, bestående av flera celler bildas. Morulan ger upphov till en vanligen ihålig blastula. Blastulan utvecklas till en tvåskiktad gastrula med en öppning, urmunnen.
Reptilerna, inklusive fåglarna, och kloakdjuren är några av flera undantag bland djuren. Hos dem klyvs inte ägget fullständigt. Endast ett litet område på äggets yta delas upp i celler och bildar en så kallad groddskiva. Detta beror på att dessa ägg har ett extremt stort förråd av upplagsnäring, gula, i form av så kallade gulekorn. Det är endast groddskivan som ger upphov till embryot. Det är bara embryot som utvecklas till den unge som kläcks ur ägget.
Människans tidiga embryonala utveckling
Efter ett dygn har befruktningen skett och vi ser två cellkärnor (pronuclei) inuti äggcellen, en hanlig (från en spermie) och en honlig (tillhörande äggcellen). Båda har ett halverat kromomantal. Äggcellen kallas nu zygot. Nedtill utanför zygoten syns en polkropp, som bildats vid den blivande äggcellens meios (reduktionsdelning). Meiosen omfattar två celldelningar. Kromosomtalet halveras under den första av dessa. Äggcellen behåller emellertid nästan all sin cytoplasma. De andra dottercellerna kallas polkroppar. De är mycket små och belägna utanför äggcellens membran.
Under det andra dygnet smälter de båda pronuclei ihop samtidigt som zygoten delar sig och ger upphov till ett embryo med två celler. Dessa celler har det normala kromosomantalet i sina kärnor, med hälften av kromosomerna från fadern och hälften från modern. Embryot genomgår sedan två celldelningar till och består nu av åtta celler. Efter ytterligare celldelningar bildar det en cellklump som kallas morula.
Morulan omvandlas efter ännu fler celldelningar till en klotformad, ihålig blastocyst. Det cellskikt som omger den kallas trofoblast och ger så småningom upphov till den yttersta fosterhinnan, korion. Innanför trofoblasten (upptill till vänster på videon) finns en cellklump, den inre cellmassan, som senare bildar en groddskiva. Det egentliga embryot bildas i groddskivan och utvecklas så småningom till ett foster. Groddskivan utvecklas i fortsättningen på i princip samma sätt hos däggdjur som hos reptiler, inklusive fåglar. Läs om fosterhinnorna och om ryggsträngen på andra sidor. Förloppen visas betydligt snabbare än de verkligen sker.
Placentadäggdjurens embryonala utveckling
Hos placentadäggdjuren försörjs ju embryot med näring från modern via moderkakan (placentan). Då behövs inte så mycket gula. Äggen har därför förlorat större delen av gulan och är gulefattiga. Märkligt nog har de i samband med detta återgått till den ursprungliga typen av klyvningsdelningar. Hela ägget delas upp i celler så att en morula uppstår. Men om du studerar den fortsatta utvecklingen, framstår det ändå som fullständigt klart, att vi placentadäggdjur härstammar från äggläggande förfäder med gulerika ägg. Den så kallade inre cellmassan i embryot delas hos oss upp i två skikt, groddblad. Det övre ektodermet ger bland annat upphov till överhud och nervsystem, det undre endodermet ger bland annat upphov till magtarmkanalens innersta cellskikt och dess körtlar. Här finns stora likheter med utvecklingen hos de äggläggande släktingarna. Ännu större likheter finner du i nästa steg, gastrulationen. Precis som hos de äggläggande släktingarna bildas en så kallad primitivstrimma genom vilken celler vandrar in mellan ektoderm och endoderm och bildar det tredje groddbladet, mesodermet.
De följande stegen i utvecklingen sker på likartat sätt hos alla ryggradsdjur. Mesodermet bildar i embryots mittlinje den så kallade ryggsträngen (kordan). På båda sidor om ryggsträngen bildas rader av somiter. Somiterna är klossar av mesodermceller. De ger bland annat upphov till läderhud och skelettmuskulator. Senare snörps ett nervrör av från ektodermet på ryggsidan ovanför ryggsträngen. De inre delarna av somiterna utvecklas till ryggkotor, som omsluter kordan och nervröret.
Placentadäggdjurens förfäder hade gulerika ägg
Primitivstrimmorna i ett människoembryo och ett hönsembryo ser nästan precis likadana ut och fungerar på samma sätt. Detta är ett starkt argument för att placentadäggdjuren och pungdjuren härstammar från djur med gulerika ägg och groddskiva. Ett andra starkt argument är att placentadäggdjuren och pungdjuren utvecklar precis samma fosterhinnor som kräldjur, inklusive fåglar, och kloakdjur. Ett tredje starkt argument är naturligtvis att kloakdjuren lägger gulerika ägg med groddskiva, men ändå har en rad egenskaper gemensamma med övriga däggdjur, till exempel hår och mjölkkörtlar. Och ett fjärde starkt argument är att icke fungerande rester av de gener som kodar för vitellogeninerna finns hos både pungdjur och placentadäggdjur. Vitellogeninerna är lipoproteiner och utgör den viktigaste komponenten i gulans näringsförråd hos de äggläggande reptilerna, inklusive fåglarna. Till yttermera visso har näbbdjuret, ett äggläggande kloakdjur, behållit en fungerande vitellogeningen.
Det fylotypiska stadiet och timglasmodellen
Det så kallade fylotypiska stadiet är en period under ryggradsdjurens embryonalutveckling, då de uppvisar stora likheter. Detta stadium infaller efter nervrörsbildningen. Embryot är då uppdelat i segment från framänden till bakänden och flera olika organ har bildats. Stadiet kallas också faryngula, eftersom gälfickor och gälbågar då har utvecklats i svalget (pharynx).
Det är mycket viktigt att notera att faryngulorna hos olika ryggradsdjur inte ser exakt likadana ut. En berömd bild publicerad av Ernst Haeckel och andra på 1800-talet är i flera avseenden felaktig och visar bland annat alltför stora likheter mellan olika djurs embryon. Biologer är sedan länge medvetna om detta, se bildtexten överst på sidan. Icke desto mindre gör kreationister fortfarande ett stort nummer av denna felaktiga bild som publicerades för mer än hundra år sedan.
Enligt den så kallade timglasmodellen finns det restriktioner under det fylotypiska stadiet, alltså hos faryngulan, som gör att alla ryggradsdjursembryon då måste vara uppbyggda efter en liknande grundplan. Utvecklingen före och efter det fylotypiska stadiet är mer flexibel. Därför finns det stora skillnader mellan olika grupper av ryggradsdjur under deras tidiga och deras sena utvecklingsstadier. Det fylotypiska stadiet motsvarar timglasets insnörning. Ett exempel är de stora skillnaderna mellan den tidiga utvecklingen hos däggdjuren och hos andra fyrfotadjur som beskrivits ovan. Den senare utvecklingen, efter faryngulastadiet, skiljer sig ju också åt hos olika ryggradsdjur. Slutresultaten blir, som vi alla vet, mycket olika varandra: en fisk, en fågel eller ett däggdjur.
Timglasmodellen stöds således av de iakttagelser som forskare kunnat göra av embryonas utveckling hos olika ryggradsdjur. Intressant nog stöds modellen också av molekylärbiologiska studier av genaktiviteterna under embryonas utveckling. Genuttrycksmönstret hos olika djur är mycket likartat under det fylotypiska stadiet. Detta innebär att det är i stort sett samma gener som är aktiva hos de olika djuren. Skillnaderna i genaktiviteter mellan olika djurgrupper är mycket större före och efter det fylotypiska stadiet.
Vad är det då som begränsar utrymmet för förändringar under det fylotypiska stadiet? Detta är fortfarande oklart, men det finns flera hypoteser. Enligt en hypotes är det nätverk av signaler, som styr utvecklingen under det fylotypiska stadiet, så komplicerat, att även små förändringar leder till kraftiga störningar. En förändrad signal kan till exempel leda till effekter i flera olika delar av embryot. Signalerna förmedlas bland annat av proteinfaktorer. Enligt en annan hypotes måste de så kallade Hox-generna uttryckas efter ett koordinerat mönster under det fylotypiska stadiet, annars utvecklas kroppens grundplan inte på ett normalt sätt.
Fördjupning
Varifrån kommer vi? Om lansettfiskar, ryggradsdjur och människans ursprung. Du hittar artikeln längre ner under den här rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
Referenser
- D. Brawand, W. Wahli och H. Kaessmann: Loss of egg yolk genes in mammals and the origin of lactation and placentation (PLoS Biology 6: e63, 2008).
- A. Collazo: Developmental variation, homology, and the pharyngula stage (Systematic Biology 49:3–18, 2000).
- N. Irie och S. Kuratani: Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis (Nature Communications 2, Article number: 248, doi:10.1038/ncomms1248, 2011).
- A. T. Kalinka et al.: Gene expression divergence recapitulates the developmental hourglass model (Nature 468:811-814, 2010).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- B. Piasecka et al.: The hourglass and the early conservation models - co-existing patterns of developmental constraints in vertebrates (PLoS Genetics 9: e1003476, 2013).
- M. K. Richardson och G. Keuck: Haeckel's ABC of evolution and development (Biological Reviews of the Cambridge Philosophical Society 77:495-528, 2002).
- W. Shumway och F. B. Adamstone: Vertebrate embryology (5th ed, Wiley, 1962).
- F. Wilt and S. Hake: Principles of developmental biology (W.W. Norton & Company, 2004).

Är människan fortfarande föremål för evolution genom naturligt urval? Det har hävdats att hon inte är det, framför allt inte i världens rika länder. Men forskare har också påvisat genetiska förändringar hos människan i förhållandevis sen tid. Hur kommer framtiden att se ut?
Har människans evolution avstannat?
Det finns flera argument som tyder på att vår evolution avstannat eller att vi inte längre är föremål för naturligt urval. En anledning skulle kunna vara att dödligheten i de rika länderna är mycket låg under tiden från och med födelsen till och med de åldrar, då vi fortplantar oss. En annan anledning skulle kunna vara att det finns en rad kulturella faktorer som påverkar antalet barn som vi sätter till världen, förutom den evolutionära fortplantningsdugligheten ("fitness"). De flesta väljer ju hur många barn de vill ha. Ytterligare en anledning skulle kunna vara att det kulturella arvet har större betydelse än det genetiska. Det kulturella arvet har tidigare inkluderat sådana saker som att lära sig att tillverka stenyxor, att försvara sig mot rovdjur och att lagra föda inför sämre tider. Numera inkluderar det bland annat skydd mot dödliga infektioner genom avloppsteknik och andra hygieniska förbättringar. drastiskt minskad barnadödlighet, framgångsrik medicinsk bekämpning av sjukdomar genom vaccin och läkemedel samt frånvaro av svält i större delen av världen. Därmed skulle det naturliga urvalet ha liten effekt på oss människor.

Bevis för evolution i geologiskt sett sen tid
Men det finns data som tyder på att vi varit föremål för naturligt urval också efter det att vi utvecklat en avancerad kultur och även fortfarande. Den ljusa hudfärgen i nordliga delar av världen utvecklades relativt sent, i Europa kanske för mindre 10 000 år sedan. Den evolutionära fördelen med ljus hudfärg har mycket sannolikt med vitamin D att göra. Storleken på våra tänder har minskat under de senaste 10 000 åren. Det tros bero på att tillagad mat är lättare att tugga än rå.
Flera folkgrupper, bland annat vi nordeuropéer, har genom naturligt urval utvecklat förmågan att dricka mjölk som vuxna. Det handlade i Europa om en mutation i en styrande gen, som i sin tur gör att genen för enzymet laktas inte stängs av, när barn slutat dia. Därmed kunde vi bryta ned sockret laktas i mjölk och drabbades inte av diarréer och andra problem vid mjölkdrickande. Detta gav boskapsskötande folkgrupper ökad tillgång till en viktig näringskälla. I Europa etablerades denna mutation kanske för mindre än 10 000 år sedan. Andra mutationer med samma effekt har uppkommit åtminstone fyra gånger i andra delar av världen.
I Afrika förekommer en genmutation som sannolikt fortfarande är föremål för naturligt urval. Den ger i dubbel dos upphov till bland annat allvarlig blodbrist, sickelcellanemi. Men har du en normal gen och en sickelcellgen är du frisk och har ökad motståndskraft mot malaria. Denna fördel gör att den muterade genen har gynnats genom naturligt urval, trots att den gör en del personer allvarligt sjuka.
Tibetanerna har genetiskt anpassats till att leva i syrgasfattig luft på mycket hög höjd, troligen efter att ha invandrat till den tibetanska högplatån för cirka 20 000 år sedan. Syrgashalten är cirka 40 procent lägre där än vid havsytan. Andra människor som kommer upp på hög höjd acklimatiserar sig inom några veckor. Bland annat ökar de halten av röda blodkroppar i blodet med hjälp av hormonet erytropoetin (EPO), så att denna halt blir högre än normalt . Då ökar blodets koncentration av det syretransporterande hemoglobinet och därmed dess syretransporterande förmåga. Funktionen med detta är egentligen att kompensera för vissa former av anemi ("blodbrist"), särskilt stora blodförluster. På hög höjd är det långsiktigt en nackdel. När halten av röda blodkroppar är högre än normalt, ökar blodets viskositet, vilket innebär att det blir mera trögflytande. Detta ökar resistansen ("motståndet") mot blodflöde i kärlen och därmed också risken för att drabbas av hjärt-kärlsjukdom. Hos tibetanerna har forskare påvisat flera genvarianter, som etablerats genom naturligt urval och som gör dem genetiskt anpassade till den syrgasfattiga miljön. En av dessa genvarianter sänker halten av röda blodkroppar, så att blodet inte blir alltför trögflytande. I stället ökar de syretillförseln till kroppen bland annat genom att ha en förhöjd viloandning och genom att ha mycket täta kapillärnät i kroppens vävnader. Märkligt nog härstammar en av tibetanernas genvarianter från denisovanerna, en utdöd människogrupp besläktad med neandertalarna.
Pågår människans evolution nu i vår tid?
Det finns många som anser att människor i sen tid har varit och fortfarande är föremål för naturligt urval. Men det är svårt att visa, bland annat på grund av vår generationslängd. Hos oss är det kanske 20–25 år eller mer mellan generationerna. Det gör att det kan ta hundratals eller tusentals år för en genmutation att etablera sig genom naturligt urval. Dessutom måste forskarna studera flera generationer för att säkert kunna påvisa evolutionär förändring. Bakterier och bananflugor har mycket korta generationstider. Hos dem kan forskarna påvisa evolutionära förändringar inom loppet av några månader eller år. Ett annat problem är att vi inte kan förutsäga framtida evolution. Evolutionen är inte riktad. Den orsakas av miljöförändringar, slumpvisa mutationer, naturligt urval och sexuellt urval.
Sexuellt urval har jag hittills inte diskuterat här. Men det finns undersökningar som tyder på att vårt val av partner inte bara styrs av kulturella faktorer, utan även av så kallat sexuellt urval. Ett exempel rör MHC-generna, som är av stor betydelse för immunförsvaret. Det finns studier som tyder på att kvinnor är mer attraherade av män med en MHC-uppsättning som skiljer sig från deras egen. Möjligen påverkas även mäns partnerval av kvinnors MHC-uppsättning, men en noggrann studie från 2017 talar emot detta. Det naturliga urvalet kan tänkas gynna ett val av en partner med annorlunda MHC-uppsättning, dels på grund av att avkomman skulle få ett effektivare immunsystem och dels på grund av att risken för inavel minskar. Mer forskning behövs dock inom detta område.
För att sammanfatta, så har människan i sen tid varit föremål för evolution med naturligt urval. Det är fullt möjligt att hon fortfarande är det, i synnerhet i utvecklingsländer. Det finns flera studier som stöder denna tanke. Det är föga sannolikt att vår evolution helt har upphört. Den sannolikaste hypotesen är att vi fortfarande undergår evolution med naturligt urval, men att evolutionen numera bara påverkar vissa egenskaper och att den inte kommer att leda till att vi drastiskt förändras eller kommer att uppdelas i olika arter.
Genvarianter som ger motståndskraft mot olika sjukdomar är troligen fortfarande föremål för naturligt urval. Om vi drabbas av en framtida global pandemi orsakad av ett virus eller en bakterie, skulle det naturliga urvalet sannolikt sortera bort de stackars människor som har en genetiskt betingad dålig motståndskraft mot smittämnet. En mycket stor studie från 2017 antyder att det naturliga urvalet nu gynnar några genvarianter, som ökar livslängden. Ett problem med detta är att äldre personer inte fortplantar sig. En av flera möjliga tolkningar av dessa resultat är att de stöder "the grandmother hypothesis".
Slutligen, även om vi fortfarande är föremål för evolution med naturligt urval, så kan vi inte förutsäga dess riktning.
Referenser
- A. Courtiol et al.: Natural and sexual selection in a monogamous historical human population (Proceedings of the National Academy of Sciences 109:8044-8049, 2012).
- A. Curry: The milk revolution (Nature 500:20-22, 2013).
- J. Hawks: Still evolving after all these years (Scientific American 311:86-91, 2014).
- M. Milinski, I. Croy, T. Hummel, och T. Boehm: Major histocompatibility complex peptide ligands as olfactory cues in human body odour assessment (Proceedings of the Royal Society B 280: 20122889, 2013).
- J. Hawks: Still evolving after all these years (Scientific American 311:86-91, 2014).
- H. Mostafavi et al.: Identifying genetic variants that affect viability in large cohorts (PLoS Biol 15[9]:e2002458, 2017).
- M. F. Small: Are human beings still evolving? It would seem that evolution is impossible now that the ability to reproduce is essentially universally available. Are we nevertheless changing as a species? (Scientific American, 1999).
- F. Probst et al.: Men’s preferences for women’s body odours are not associated with human leucocyte antigen (Proceedings of the Royal Society B 284:20171830, 2017).
- F. M. Salzano: Is the human species still evolving? To where? (Genetics and Molecular Biology 35:899-903, 2012).
- J. T. Stock: Are humans still evolving? (EMBO reports 9:S51-S54, 2008).
- P. Tyson: Are We Still Evolving? (Nova, 2009).
- J. C. Winternitz och J. L. Abbate: Examining the evidence for major histocompatibility complex-dependent mate selection in humans and nonhuman primates (Research and Reports in Biology 6:73-88, 2015).

Vilken är människans äldsta kända förfader? Vilket nu levande djur är mest likt denna förfader? De äldsta fossil som vi känner till i vår utvecklingshistoria är cirka 500 miljoner år gamla. Fantastiskt nog visar det sig att dessa fossil är mycket lika en nu levande djurgrupp, lansettfiskarna, så till den grad att lansettfiskarna kan betraktas som levande fossil. De är, trots sitt namn, inte fiskar.
Människan är ett däggdjur. Däggdjuren tillhör understammen ryggradsdjur (Vertebrata) inom stammen ryggsträngsdjur (Chordata) bland djuren. Till ryggradsdjuren hör alla fiskar och alla fyrfotadjur (det vill säga groddjur, kräldjur inklusive fåglar, och däggdjur). Ryggsträngsdjuren inkluderar ytterligare två understammar: manteldjur (Urochordata; se bilden nedan) och lansettfiskar (Cephalochordata, se bilden ovan). Manteldjuren och lansettfiskarna har en betydligt enklare kroppsbyggnad än ryggradsdjuren. De har därför sedan gammalt ansetts stå närmare ryggsträngsdjurens urfader än ryggradsdjuren. Detta har nu bekräftats av molekylärbiologiska studier av arvsmassans DNA.
Ryggsträngsdjurens urfader levde förmodligen långt innan den så kallade "kambriska explosionen", som inleddes för cirka 540 miljoner år sedan. Notera att Pikaia, det ursprungligaste kända fossila ryggsträngsdjuret, levde betydligt senare, för cirka 505 miljoner år sedan. Se bilden överst på sidan. Under den kambriska explosionen uppträdde plötsligt de flesta av de artrikaste nu levande djurstammarna på den fossila arenan. Orsakerna till detta är oklara. Det finns emellertid ett flertal hypoteser. Molekylärbiologiska data tyder på att det kanske inte var någon explosion. I stället uppkom kanske flera av djurgrupperna långt innan kambriumperioden började. Orsaken till att forskarna har hittat så få fossil från tiden före kambrium skulle kunna vara djuren då saknade mineraliserade skelett och skal. Mjukdelar bevaras mycket sällan som fossil.
Lansettfiskarna är de nutida djur som står närmast ryggsträngsdjurens urfader, men de är inte ryggradsdjurens närmaste nu levande släktingar. Förvånande nog tyder DNA-studier i stället på att det är manteldjuren som står oss ryggradsdjur närmast. Du kan ju knappast tänka sig ett djur som är mer olikt ett ryggradsdjur än manteldjuret på den översta bilden nedan, en vuxen sjöpung. Förklaringen är att manteldjurens larver ser helt annorlunda ut än de vuxna djuren och har en rad anatomiska egenskaper som är gemensamma för alla ryggsträngsdjur. Mer om dessa egenskaper senare. Som synes på den nedersta bilden nedan, påminner manteldjurens larver till det yttre om grodlarver. De genomgår en drastisk omvandling, en metamorfos, som resulterar i ett vuxet djur med en radikalt annorlunda kroppsbyggnad. En grupp bland manteldjuren behåller dock flera larvala egenskaper som vuxna.

Genomfördubblingar och Hox-gener
Lansettfiskarna och manteldjuren har en mycket enkelt uppbyggd kropp. Det samma gällde Pikaia, det ursprungligaste kända fossila ryggsträngsdjuret, som levde för cirka 505 miljoner år sedan. Ryggradsdjurens förfader påminde med största sannolikhet om Pikaia och lansettfiskarna. Hur kunde ett så enkelt uppbyggt djur, utan hjärna och hjärta, ha gett upphov till den utvecklingslinje, som ledde fram till dagens komplicerat uppbyggda ryggradsdjur, bland dem människan? Förklaringen finner du i en genomgripande händelse som ägde rum för mer än 500 miljoner år sedan. Vid två tillfällen fördubblades hela genomet (arvsmassan) hos den utvecklingslinje bland ryggsträngsdjuren, som senare gav upphov till ryggradsdjuren. Forskarna tror numera att detta skedde hos en urfader till alla ryggradsdjur. Men den andra genomfördubblingen kan också ha skett efter det att de käklösa ryggradsdjuren, rundmunnarna (Agnatha), grenades av från släktträdet, men innan de käkförsedda ryggradsdjuren (Gnathostomata) avgrenades. Till rundmunnarna hör i dag bara pirålar och nejonögon. Alla övriga ryggradsdjur är käkförsedda. De inkluderar alla fiskar, utom pirålar och nejonögon, samt alla fyrfotadjur, det vill säga groddjur, kräldjur inklusive fåglar och däggdjur. Om två genomfördubblingar inte hade skett, så hade det inte funnits några ryggradsdjur på jorden, utom möjligen pirålar och nejonögon med en genomfördubbling.
Två genomfördubblingar ledde till att det i princip fanns fyra gener i stället för en av alla genomets gener, egentligen fyra par av varje gen fördelade på fyra kromosompar. Nästan alla kroppscellernas gener hos djur finns nämligen normalt i två exemplar, ett på vardera kromosomen i ett kromosompar. Det fanns nu tre nya genkopior, som inte var nödvändiga. De flesta av de nya genkopiorna hos urryggradsdjuren försvann emellertid så småningom från genomet. Andra drabbades av skadliga mutationer, som ledde till att de inte fungerade längre. De blev då genetisk barlast, så kallade pseudogener. Men många nya genkopior fortsatte att fungera. Vi kan fråga sig hur avkomlingar, försedda med ett ökat antal identiska genkopior, kunde utveckla en kropp med ett mera komplicerat byggnadsplan. Förklaringen är att många av de nya genkopiorna under inflytande av naturligt urval genomgick en evolution så att de fick förändrade funktioner jämfört med den ursprungliga genen. Särskilt betydelse hade detta för de så kallade homeotiska generna, som styr uppkomsten av kroppens byggnadsplan under embryots utveckling. De mest kända bland de homeotiska generna är Hox-generna, som hos djur styr utvecklingen av kroppens delar i dess längsriktning, hos däggdjur från huvudet till svansen. Hox-generna ligger vanligen intill varandra längs med en kromosom i samma ordningsföljd som de kroppsdelar de påverkar.

Lansettfiskarna har en uppsättning Hox-gener. Ryggradsdjuren har fyra uppsättningar Hox-gener. De har emellertid förlorat några Hox-gener. Därför har de inte fyra gånger fler Hox-gener än lansettfiskarna. Men de har avsevärt fler Hox-gener med olika funktioner än lansettfiskarna. De har också fler homeotiska gener med andra funktioner än Hox-generna. Ett ökat antal homeotiska gener med nya funktioner är den sannolikt viktigaste orsaken till att ryggradsdjuren har kunnat utveckla en avsevärt mera komplicerad kroppsbyggnad än lansettfiskarna och även flera nya organ.
Ryggsträngsdjurens utveckling
Ryggsträngsdjuren förenas av ett antal gemensamma anatomiska strukturer. Hos många av dem uppträder dessa strukturer bara under embryots utveckling i ägget, hos en del också under larvstadiet, men de finns där.
Ryggsträngen (kordan) och ryggraden
Ryggsträngen är den struktur som gett namn åt djurgruppen. Den kallas även korda. Kordan är en elastisk stav som löper längs med ryggen och fungerar som skelett. Den innehåller en kärna av celler och utanför dem vätska, ibland finns vätskan inuti cellerna. Utanför kärnan finns ett segt hölje av bindvävnad. Eftersom vätskan inte kan pressas ihop, kan kordan inte förkortas. När muskulaturen på ena sidan av kroppen dras ihop, böjs I stället kordan, och därmed hela kroppen, i en båge. När muskulaturen slappnar av fjädrar den elastiska kordan tillbaka, så att kroppen åter blir rak. Detta gör att djuret kan simma på ett effektivt sätt. Kordan fungerar som ett så kallat hydrostatiskt skelett.

En välutvecklad korda finns hos de vuxna lansettfiskarna och hos manteldjurens larver. De flesta manteldjur saknar korda som vuxna. Ryggradsdjuren kan, som ovan nämnts, indelas i två grupper. Till rundmunnarna (Agnatha) hör pirålar och nejonögon. De har inte utvecklat käkar. De övriga ryggradsdjuren (Gnathostomata) har käkar. Rundmunnarna har en välutvecklad korda även som vuxna. Hos pirålarna utgörs ryggskelettet enbart av en korda. Nejonögonen har dessutom ryggkotor. Hos de käkförsedda ryggradsdjuren (Gnathostomata) har ryggraden med sina kotor av brosk eller benvävnad i regel tagit över kordans funktioner. Kordan finns dock kvar hos en del vuxna fiskar, bland annat hos lungfiskar och störfiskar. Hos människan utgör den geléartade kärnan i de mellan ryggkotorna belägna diskarna en rest av kordan. Flera anatomiska egenheter hos pirålarna, bland annat frånvaron av ryggrad, gör att forskarna ibland räknar dem som en systergrupp till ryggradsdjuren inom den överordnade gruppen Craniata ("djur med kranier"). Denna indelning stöds emellertid inte av molekylärbiologiska data.
Nervröret och nervsystemet
Ett andra kännetecken för ryggsträngsdjuren är nervsystemet anläggs längs med embryots hela ryggsida som en långsträckt inbuktning av ektodermet, embryots yttersta cellskikt, som också ger upphov till överhuden. Inbuktningen fördjupas och snörps av från huden som ett nervrör. Hos vuxna lansettfiskar finns nervröret kvar med en uppsvällning i framändan som tros vara ett förstadium till en hjärna. Hos ryggradsdjuren omvandlas det enkla nervröret under embryots utveckling till deras komplicerade nervsystem med hjärna och ryggmärg.
Nederst ses undre delen av ett tvärsnitt genom en lansettfisk, bakom framänden. Du ser gältarmen med en mängd gälbågar och mellan dem öppningar, gälspringor. Precis som manteldjuren, intar lansettfiskarna sin föda genom att filtrera bort plankton och organiska partiklar från havsvattnet. Vattnet tas in genom munnen, varvid grövre partiklar filtreras bort av munapparaten. Det passerar sedan ut genom gälspringorna. Då fastnar ätbara partiklar på de slemklädda gälbågarna. Slemmet med de matnyttiga partiklarna transporteras sedan av cellernas flimmerhår (cilier) uppåt längs med gälbågarna och sedan bakåt längs med en fåra i gältarmens tak. Till slut når slemmet den egentliga tarmen, bakom gältarmen. Där bryts födan ner och nedbrytningsprodukterna absorberas till blodet. Vattnet som lämnar gältarmen hamnar i atrium, ett särskilt hålrum som släpper ut vattnet genom en atriopor på djurets undersida.
I gältarmens botten finns hypostylen, som är en föregångare till ryggradsdjurens sköldkörtel. En blindsäck (cecum) som utgår från tarmen bakom gältarmen, anses vara en föregångare till ryggradsdjurens lever. Den försörjs nämligen, precis som levern, av en så kallad portåder, genom vilken den tillförs blod direkt från tarmens kapillärsystem. Notera på den övre bilden att gälbågar och gälspringor är snedställda. Det är därför som de syns i tvärsnitt på den undre bilden. Färgade ljusmikroskopiska snitt.
Gältarmen, gälbågarna och käkarna
Ett tredje kännetecken för ryggsträngsdjuren är gältarmen. Gältarmen är en svalgregion försedd med tvärgående springor, gälspringor, som avgränsas av gälbågar. Gältarmens ursprungliga funktion var sannolikt att filtrera födopartiklar från vattnet. Denna funktion har den fortfarande hos manteldjur och lansettfiskar. Eftersom den genomspolas av vatten, kan den samtidigt fungera som andningsorgan.
Manteldjur och lansettfiskar har ett mycket stort antal gälbågar. De första ryggradsdjuren var fiskar. Hos dem reducerades antalet gälbågar. De återstående gälbågarna försågs med skelett av brosk eller ben. Samtidigt försågs gältarmen med muskulatur och gälbågarna med ytförstorade utskott, gälar. Gältarmen fungerade därmed som ett andningsorgan med en muskeldriven pump, inte som en filtreringsapparat. Detta utvecklingsstadium finner vi i princip hos de nutida käklösa ryggradsdjuren, pirålar och nejonögon, även om deras gälapparat är starkt modifierad.
Hos de käkförsedda ryggradsdjuren omvandlades först den främsta gälbågen, mandibularbågen, eventuellt flera främre gälbågar, till käkar med käkleder. Nästa gälbåge, hyoidbågen, utvecklades sedan till ett stöd för käkarna. De bakre fem gälbågarna bar gälar och ingick i andningsapparaten. Detta utvecklingsstadium finner vi i princip hos alla nutida fiskar, utom pirålar och nejonögon. Uppkomsten av käkar var en revolution som gjorde det möjligt att utnyttja många nya födokällor.
Hos de fyrfota ryggradsdjuren modifierades käkarna, allra mest hos däggdjuren. Hos däggdjuren har den gamla käkleden, fantastiskt nog, ryckt in i mellanörat och en ny käkled utbildats. Däggdjurens tungben (hyoidben) härstammar till största delen från hyoidbågen. Det är det enda större ben som saknar förbindelse med det övriga skelettet. Hos fyrfotadjuren uppträder gälbågarna under embryots utveckling och ger upphov till flera strukturer i halsregionen, men de tillbakabildas sedan.
Hypostylen och sköldkörteln
Ett fjärde kännetecken för ryggsträngsdjuren är att de insöndrar jodhaltiga hormoner från särskilda celler. Hos lansettfiskar och manteldjur insöndras dessa hormoner från celler i hypostylen i gältarmens botten, se bilden ovan. Hos ryggradsdjuren bildar dessa celler en särskild körtel, sköldkörteln, som hos människan är belägen på halsen framsida. Bisköldkörtlarna härstammar från två par gälbågar. Det är därför som de är fyra stycken, ett par från varje gälbåge.

Neurallist och plakoder
Två embryonala nybildningar har haft oerhört stor betydelse för ryggradsdjurens utveckling: neurallisten och plakoderna. Precis som nervröret, härstammar dessa strukturer från ektodermet, embryots yttersta cellskikt. Neurallisten utgörs av embryonala celler som uppkommer i det område, där nervröret snörps av. Dessa celler vandrar ut i hela embryot och utvecklas till en rad viktiga vävnader och celler. Lansettfiskarna bildar ingen neurallist. Manteldjuren gör det, men deras list ger bara upphov till pigmentceller. Plakoderna är förtjockningar av ektodermet. De ge bland annat upphov till luktorganen, ögonens linser samt inneröronens balansorgan inklusive hörselorganen. Lansettfiskar och manteldjur saknar plakoder. Haikouella och andra fossil från kambriumperioden har tolkats som föregångare till ryggradsdjuren eller tidiga ryggradsdjur med neurallist, men tolkningarna är osäkra.
Fördjupning
Hur kryper daggmaskar och plattmaskar? Du hittar artikeln under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?.
Referenser
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).
- J.-Y. Chen: Early crest animals and the insight they provide into the evolutionary origin of craniates (Genesis 46:623–639, 2008).
- S. Conway Morris och J.-B. Caron: Pikaia gracilens Walcott, a stem-group chordate from the middle Cambrian of British Columbia (Biological Reviews 87:480–512, 2012).
- F. Delsuc, G. Tsagkogeorga, N. Lartillot, och H. Philippe: Additional molecular support for the new chordate phylogeny (Genesis 46:592–604, 2008).
- D. S. Fudge, N. Levy, S. Chiu, och J. M. Gosline: Composition, morphology and mechanics of hagfish slime (Journal of Experimental Biology 208:4613-4625, 2005).
- J. Garcia-Fernàndez och È. Benito-Gutiérrez: It's a long way from amphioxus: descendants of the earliest chordate (BioEssays 31:665–675, 2009).
- L. Z. Holland et al.: The amphioxus genome illuminates vertebrate origins and cephalochordate biology (Genome Research 18:1100-1111, 2008).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- Nicholas H. Putnam et al.: The amphioxus genome and the evolution of the chordate karyotype (Nature 453:1064-1072, 2008).
- N. Takatori et al.: Comprehensive survey and classification of homeobox genes in the genome of amphioxus, Branchiostoma floridae (Development, Genes and Evolution 218:579–590, 2008).

Många människor har i alla tider undrat om det finns liv i universum på andra ställen än Jorden. Särskilt har de undrat om det finns intelligent liv. Andra har, med tanke på människans historia, undrat om det finns intelligent liv på Jorden. Jag lämnar den sista frågan därhän. Här följer några funderingar om möjligheterna för utomjordiskt liv i universum.
Vi vet naturligtvis ingenting om livets uppkomst på Jorden. Fossilen ger oss ingen upplysning. Forskare har emellertid gjort många modellförsök, som pekar på rimliga mekanismer för uppkomst av liv. Den gemensamma genetiska koden hos alla organismer talar mycket starkt för att det liv som nu finns på Jorden har ett gemensamt ursprung. Förutsättningen för att nytt liv ska utvecklas finns däremot sannolikt inte på Jorden i dag. Det är mycket troligt att den speciella miljö, som fanns när den unga Jorden svalnat var särskilt gynnsam för uppkomst av liv.

Finns det okända livsformer på Jorden?
Men liv skulle mycket väl kunna ha uppstått flera gånger på Jorden. I så fall kan okända livsformer ha slagits ut av det naturliga urvalet. Det kan till och med vara så att liv med annat ursprung än det liv vi känner till, finns på Jorden i dag. Det vi i dag vet om bakterier, de bakterieliknande arkéerna och virus är bara toppen på ett isberg. Antalet kända former inom dessa grupper har ökat enormt, sedan forskare började göra storskaliga sekvenseringar av DNA bland annat från havsvatten och till och med från människans tarmar. Det måste vara oerhört svårt att hitta eventuella bakterieliknande organismer med en annan genetisk mekanism på undangömda ställen som i berggrunden och på djuphavets botten.
Drakes ekvation
Francis Drake ställde år 1961 upp en formel som ger det sannolika antalet civilisationer, som i vår galax kommunicerar med elektromagnetisk strålning upptäckbar för oss. Ekvationen innehåller följande faktorer: antalet stjärnor som bildas per tidsenhet i galaxen, andelen av dessa stjärnor som har planeter, andelen av dessa planeter som har förutsättningar för liv, andelen av de senare planeterna på vilka liv utvecklas, andelen planeter med liv på vilka civilisationer utvecklas, andelen civilisationer som sänder ut elektromagnetisk strålning och den sannolika livslängden för en sådan civilisation. Problemet är värdena på dessa faktorer är mycket osäkra eller okända. Så vi kan inte besvara frågan om det finns liv på andra planeter eller månar. Men vi kan överväga vad som är det rimligaste svaret på den frågan, med Drakes formel som utgångspunkt.
Exoplaneter, planeter utanför vårt solsystem
Under senaste två decennierna har astronomer upptäckt ett mycket stort antal så kallade exoplaneter, det vill säga planeter vid andra stjärnor än Solen. Forskarna har hittat betydligt fler än 3 000 exoplaneter. Vissa av planeterna har en del av de egenskaper som har gynnat livet på Jorden. Forskarna har dock ännu inte hittat en planet med alla de egenskaper som möjliggör liv på Jorden. Men med tanke på det enorma antalet stjärnor i galaxen, borde det vara mycket sannolikt att sådana planeter finns och att några av dem hyser liv.
Hur sannolikt är det att det finns utomjordiskt liv?
Jorden uppkom för cirka 4,5–4,6 miljarder år sedan. Fossilen av bakterieliknande celler visar att liv uppkom förvånande snabbt efter det att den unga Jorden svalnat. Detta skulle kunna tolkas som om liv uppkommer så snart en gynnsam miljö finns. Själv tror jag att det finns en mängd planeter i galaxen med bakterieliknande livsformer. Dessutom finns några månar vid Jupiter och Saturnus som tros innehålla oceaner av flytande vatten unde den fasta ytan. Många menar att liv kan ha uppkommit i dessa oceaner. Många menar också att liv kan ha funnits på planeten Mars. Så vi kan inte utesluta möjligheten att liv uppkommit flera gånger i vårt eget solsystem.
Efter livets uppkomst på Jorden dröjde det cirka 3 miljarder år, tills det att de första djuren uppträdde som fossil och ytterligare cirka 0,5 miljarder år innan den mänskliga civilisationen uppkom. Detta skulle kunna tolkas som att sannolikheten för att först komplicerat flercelligt liv och sedan civilisationer uppkommer är mindre än att enkelt liv uppkommer. Innan komplicerade livsformer uppkommit, skulle livet på en planet mycket väl kunna släckas ut av enorma vulkaniska utbrott, av gigantiska meteoriter eller av strålningspulser från närbelägna stjärnor. För cirka 250 miljoner år sedan, vid permperiodens slut, inträffade det största kända av flera stora utdöenden bland Jordens organismer. Denna händelse kunde mycket väl ha lett till att livet helt utrotats på vår planet. Orsakerna till utdöendet är omdiskuterade.
Civilisationer med avancerad teknik har antagligen en begränsad livslängd. Om de inte utplånas av katastrofer utom deras egen kontroll, skulle de mycket väl kunna förstöra den planet de lever på. Här på Jorden är vi ju på väg att göra det och måste enas om att bevara vår planet, innan det är för sent.
Hur skulle utomjordiskt liv kunna se ut?
Forskare har spekulerat mycket om hur avancerat byggda utomjordiska organismer skulle kunna se ut. Själv tror jag att vi skulle känna igen mycket av oss själva i deras uppbyggnad. Samma kemiska och fysikaliska lagar gäller ju i hela universum. Jag tror att utomjordiskt liv, precis som jordiskt, består av kolföreningar lösta i vatten. Grundämnet kol har unika egenskaper. Inget annat grundämne kan bilda så många kemiska föreningar som kol. Forskarna har dock föreslagit att liv skulle kunna baseras på grundämnet kisel. Likaså har vatten unika egenskaper som lösningsmedel för liv. Forskarna har framkastat tanken att liv baserat på flytande kolväten skulle kunna finnas på Saturnus måne Titan, men det återstår att visa. Främmande planeter är tredimensionella med tyngdkraft, precis som Jorden. Avancerat byggda djurliknande landlevande organismer där borde därför vara bilateralsymmetriska och försedda med huvud, sinnesorgan och gångben, precis som de flesta djur på Jorden. Men det kan naturligtvis också finnas liv i former, som är helt utanför vår föreställningsvärld.
Kommer vi att upptäcka utomjordiskt liv?
Med tanke på det enorma antalet stjärnor i vår galax och det enorma antalet galaxer i universum, måste den rimligaste utgångspunkten trots allt vara att det finns liv och civilisationer på planeter eller månar vid andra stjärnor än solen. Frågan om vi någonsin kommer att kunna kommunicera med andra civilisationer är betydligt svårare. På grund av de enorma avstånden mellan stjärnorna kan det synas mindre sannolikt. Organisationen SETI letar med hjälp av radioteleskop efter signaler från utomjordiska civilisationer, men har inte hittat några. En del menar att vi skulle sända meddelande ut i rymden i hopp om att de ska uppfångas av utomjordingar. Andra menar att vi absolut inte ska göra detta. De hänvisar till människans historia med de krig och folkmord som har följt efter kontakter mellan olika kulturer och folk, till exempel när västerlänningar koloniserade Amerika, Afrika och andra delar av världen.

Fördjupning
Radiär och bilateral symmetri hos djur. Tvåsidiga djur. Varför har vi höger och vänster kroppshalva? Artikeln finns under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?.
Att läsa
- J. Abbott och E. Persson (red.): LIV Utomjordiskt Syntetiskt Artificiellt (Pufendorfinstitutet, 2017).
- D. Dunér (red.): Extrema världar. Om sökandet efter liv i rymden (Pufendorfinstitutet, 2013).
- Anonymous: What Is an Exoplanet? (NASASpacePlace, uppdaterad 23 maj 2018).
- S. Seager och W. Bains: The search for signs of life on exoplanets at the interface of chemistry and planetary science (Science Advances 1(2): e1500047, 2015).
- B. Stenholm (red.): Astrobiologi (Liber, 2012).

Pleistocenepoken varade från cirka 2,6 miljoner år sedan fram till den senaste istidens slut för cirka 12 000 år sedan. Under början av denna epok fanns det i hela världen ett mycket stort antal nu utdöda stora landdjursarter, främst däggdjur, många av dem jättestora. Varför försvann alla dessa djur?
Framför allt under slutet av pleistocen dog en oerhört stor del av dessa djurarter ut på alla jordens kontinenter. I Afrika dog dock färre djurarter ut än i andra världsdelar. Utdöendet fortsatte under början av holocenepoken, efter istidens slut. Vi vet inte varför alla dessa djur dog ut, men flera hypoteser finns.

De stora djurarterna räknas till den så kallade megafaunan, som omfattar landdjur tyngre än cirka 40–45 kg. Jag ger först några exempel på utdöda megafaunadjur i olika världsdelar och diskuterar sedan möjliga orsaker till utdöendet, ett mycket kontroversiellt ämne.

Europa och Asien
I Europa och Asien försvann bland annat ullhårig mammut och flera andra elefantdjur, ullhårig noshörning, flera hästarter, jättehjort, stäppbison, flera flodhästarter, grottlejon, grottbjörn och grotthyena. Här dog också neandertalmänniskan och de så kallade denisovanerna ut.

Nordamerika, Sydamerika och Australien
I Nordamerika försvann bland annat mammutar, mastodonter (en grupp elefantdjur), jättebison och andra bisonarter, jättebäver, flera hästarter, flera arter av jättetrögdjur (släkt med dagens sengångare), flera kameldjur samt det berömda sabeltandade kattdjuret Smilodon (som även fanns i Sydamerika). Här försvann också teratornierna, som var jättestora flygande fåglar.

I Sydamerika försvann bland annat flera elefantdjur, flera jättetrögdjur, flera hästarter, flera kameldjur, teratornierna samt de jättestora glyptodonterna (släkt med dagens bältor), bland annat spikklubbsbältorna. Här dog också två hela däggdjursordningar ut, litopternerna och notoungulaterna. De var hovdjur och hade båda uppkommit i Sydamerika under den långa tid, då kontinenten var isolerad från andra världsdelar.

I Australien försvann flera gigantiska pungdjur, bland annat jättekängurur, de så kallade punglejonen och den noshörningsstora jättevombaten (Diprotodon). Där försvann också en marklevande jättefågel (Dromornis) och en jättevaran. Den senare var mycket större än komodovaranen, den största nu levande ödlan.
Orsaker till utdöendet: klimatet och människan
Orsakerna till megafaunornas utdöende är mycket omdiskuterade. De två viktigaste hypoteserna är klimatförändringar under och efter den senaste istiden och utrotning genom ingripande av den moderna människan (Homo sapiens). Det är möjligt, för att inte säga sannolikt, att det fanns flera samverkande orsaker. Det är också möjligt att det inte var samma huvudorsak i alla delar av världen.
Hela pleistocenepoken kännetecknades av drastiska klimatförändringar med flera istider och mellanliggande varmare perioder. Även under en pågående istid varierade klimatet. Dessa klimatfluktuationer åtföljdes av stora förändringar i vegetationen. En invändning mot klimathypotesen är att megafaunan till största delen överlevde alla dessa svängningar i klimatet. Varför dog då så många djurarter då ut under en relativt kort period kring den senaste istidens slut? Mot detta argument kan anföras att populationerna skulle kunna ha minskat vid varje klimatomslag, i och för sig överlevt, men med reducerat individantal. I så fall skulle uppvärmningen vid den senaste istidens slut kunna ha varit nådastöten för ett stort antal djurarter. Stora djur har svårare att anpassa sig till varmare klimat än små, eftersom de lättare drabbas av överhettning. Orsaken är att de har en mindre värmeavgivande kroppsyta räknat per kilogram kroppsvikt med värmeproducerande vävnad.

Den moderna människan, Homo sapiens, utvandrade ur Afrika under slutet av pleistocenepoken. Tidpunkten är omdiskuterad. Migrationen tros ha skett för mellan cirka 120 000 och 50 000 år sedan. Det kan mycket väl ha varit flera olika migrationer vid olika tidpunkter och via olika vägar. De olika kontinenterna befolkades också vid olika tidpunkter av moderna människor. Australien nåddes troligen för cirka 60 000–40 000 år sedan, Europa för cirka 45 000–30 000 år sedan och Amerika för cirka 14 000 år sedan, eventuellt ännu tidigare.
Vår människoart hade utvecklat en mera avancerad jaktteknologi än andra människoarter. Vi använde sannolikt kastspjut. Vi hade kanske utvecklat ett mera effektivt samarbete under jakten. Det skulle därför kunna ha varit vi som utrotade en mängd djurarter genom jakt eller genom annan miljöpåverkan. Mot denna förklaring har anförts att arkeologiska bevis för storskalig jakt är sällsynta. Dessutom sammanfaller utdöenden inte alltid med människan ankomst till ett område i världen. Å andra sidan skulle det kunna vara så att megafaunan klarade sig bättre i Afrika tack vare att den där utvecklats parallellt med olika människoarter under flera miljoner år.
Vi kan fråga oss, om det var värt mödan och risken, att med primitiva vapen angripa farliga mammutar och noshörningar, så länge vi hade tillgång till mindre bytesdjur. Å andra sidan fanns det väldigt mycket mat i en mammut.

Orsaker till utdöendet: sjukdom och kometnedslag
Det finns några andra möjliga förklaringar till massutdöendet. De flesta anser dock dessa hypoteser mindre sannolika.
Det har spekulerats att människan, när hon spridde sig till nya områden, också spridde sina infektionssjukdomar till andra djurarter. Människan hade under sin evolution utvecklat en motståndskraft mot dessa sjukdomar, men för arter som aldrig hade drabbats av smittämnena var de dödliga. Det finns inga belägg för denna hypotes. Från historisk tid känner vi dock till flera exempel på motsatsen: spridning av farliga infektioner från andra djur till människor. Influensavirus härstammar från bland annat svin och fåglar. Smittkoppsvirus anses ha överförts till människan från andra däggdjursarter, pestbakterien från gnagare och Ebolavirus från fladdermöss.
Enligt en annan hypotes bidrog ett belagt nedslag av ett utomjordiskt objekt, förmodligen en komet, för cirka 13 000 år sedan till megafaunans utdöende i Nordamerika.
Orsaker till utdöendet: sammanfattning
Vilka orsakerna till utdöendena än var, är det viktigt att framhålla att stora djur har glesare populationer och långsammare fortplantning än små djur. Stora djur är därför mycket känsligare för alla slags miljöförändringar.
År 2018 publicerades en omfattande studie som jämförde utdöenden dels bland däggdjuren från 65 miljoner år före nutid fram till 125 tusen år före nutid, dels bland däggdjuren under de senaste 125 tusen åren. Forskarna fann att de stora djuren, megafaunan, var överrepresenterade bland de djur som dog ur under de senaste 125 tusen åren, jämfört med de som dog ut tidigare. Detta gällde alla kontinenter och alla studerade däggdjursgrupper. Under dessa 125 tusen år utvandrade den moderna människan (Homo sapiens) från Afrika och befolkade först Eurasien och Australien, sedan Amerika. Forskarna fann också data som tydde på att megafauna dog ut i Afrika redan innan Homo sapiens uppträdde på arenan. Allt detta tolkades som ett stöd för hypotesen att människan starkt bidrog till megafaunans utdöende. Vi bör dock komma ihåg att det är en korrelation, som inte säger något om orsakssammanhangen. Dessutom är dateringar av utdöenden bristfålliga för många arter.
Sammanfattningsvis så vet vi för lite om megafaunans kollaps för att ännu kunna fastställa vad som orsakade den. Mycket tycks peka på att det fanns flera samverkande orsaker.

Australien, Madagaskar och Nya Zeeland: människan bar huvudansvaret
Merparten av Australiens megafauna dog sannolikt ut redan för cirka 50 till 40 tusen år sedan, i samband människans ankomst till denna världsdel. Här var kanske människans ingripande den viktigaste orsaken till utdöendet, även om ett torrare klimat kan ha bidragit. Människan ingrep inte bara genom jakt, utan troligen också genom att bränna vegetationen inom stora områden.
Madagaskar befolkades i relativt sen tid, från Sydostasien och Afrika med början för cirka 2 300 år sedan. De gigantiska elefantfåglarna och de stora lemurerna på Madagaskar utrotades sannolikt av människan. Eventuellt bidrog även klimatförändringar till deras försvinnande.

Nya Zeeland var den sista större landmassa som befolkades. Detta hände för cirka 700 år sedan. Forskarna är tämligen säker på att det där var människan som utrotade megafaunan. Den bestod av fåglar. Det fanns många arter av strutsliknande moafåglar, flera av dem mycket stora. Det fanns också en jättestor örn som tros ha dött ut när dess byte, moafåglarna, försvann.
Nutidens utdöende: en potentiell katastrof
Det stora utdöende som pågår i vår tid är utan tvekan orsakat av människan, framför allt genom direkt miljöpåverkan men även jakt och fiske. Det är jämförbart med udöendet vid kritaperiodens slut för cirka 70 miljoner år sedan, då bland annat dinosaurierna utom fåglarna dog ut. Det är till och med jämförbart med utdöendet vid permperiodens slut för cirka 250 miljoner år sedan, då djurlivet på Jorden nästan utplånades. Det är människans ansvar att se till att en sådan katastrof inte händer i framtiden.
Fördjupning
Vilken nytta har djur av sin svans? Varför har människan förlorat svansen? Du hittar artikeln under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?.
Världens största ödla, komodovaranen. Artikeln finns under rubriken Märkliga och farliga djur.
Referenser
- M. E. Allentoft et al.: Extinct New Zealand megafauna were not in decline before human colonization (Proceedings of the National Academy of Science 111:4922-4927, 2014).
- The art of Altamira (Museo Nacional y Centro de Investigación de Altamira, hämtad 9 november 2017).
- Australia's extinct animals (Australian Museum, hämtad 16 november 2017).
- M. Bunce et al.: Ancient DNA provides new insights into the evolutionary history of New Zealand's extinct giant eagle (PLoS Biology 3:e9, 2005).
- B. E. Crowley: A refined chronology of prehistoric Madagascar and the demise of the megafauna (Quaternary Science Reviews 29:2591-2603, 2010).
- Encyclopedia of Life (hämtad 6–16 november 2017).
- R. B. Firestone et al.: Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and the Younger Dryas cooling (Proceedings of the National Academy of Science 104:16016-16021, 2007).
- J. L. Gill et al.: Pleistocene megafaunal collapse, novel plant communities, and enhanced fire regimes in North America (Science 326:1100-1103, 2009).
- C. V. Haynes et al.: The Murray Springs Clovis site, Pleistocene extinction, and the question of extraterrestrial impact (Proceedings of the National Academy of Science 107:4010-4015, 2010).
- C. Johnson: Megafaunal decline and fall (Science 326:1072-1073, 2009).
- J. L. Metcalf et al.: Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation (Science Advances 2: e1501682, 2016).
- Nationalencyklopedin (nedladdat 6–16 november 2017).
- S. R. Pant, A. Goswami, och J. A Finarelli: Complex body size trends in the evolution of sloths (Xenarthra: Pilosa) (BMC Evolutionary Biology 14:184, 2014).
- J. L. Prado, C. Martinez-Maza, och M. T. Alberdi: Megafauna extinction in South America: A new chronology for the Argentine Pampas (Palaeogeography, Palaeoclimatology, Palaeoecology 425:41-49, 2015).
- N. J. Rawlence et al.: Soft-tissue specimens from pre-European extinct birds of New Zealand (Journal of the Royal Society of New Zealand 43:154-181, 2013).
- R. G. Roberts och B. W. Brook: And then there were none? (Science 327:420-422, 2010).
- N. Rust: What really happened to mammoths and other ice age giants (BBC Earth, uppdaterad 2 november 2016).
- Freédérik Saltré et al.: Climate change not to blame for late Quaternary megafauna extinctions in Australia (Nature Communications 7:10511, 2016).
- C. Sedwick: What killed the woolly mammoth? (PLoS Biology 6:e99, 2008).
- F. A. Smith et al.: Body size downgrading of mammals over the late Quaternary (Science 360:310-313, 2018).
- B. Switek: What killed South America's megamammals (Scientific American Blog Network, uppdaterad 21 juni 2016).
- E. P. Tonni, A. L. Cione, L. H. Soibelzon: The broken zig-zag: Late Cenozoic large mammal and tortoise extintion in South America (Revista del Museo Argentino de Ciencias Naturale 5:1-19, 2003).

Nästa alla djur, växter och svampar fortplantar sig könligt med hjälp av hanliga och honliga könsceller, det vill säga spermieceller och äggceller, eller med hjälp av hanliga och honliga cellkärnor. Detsamma gäller merparten av de encelliga kärnförsedda organismer som kallas protister. Många arter inom dessa organismgrupper kan dessutom fortplanta sig på könlös väg, till exempel genom avknoppning av kroppsdelar eller genom celldelning. Men det är mycket få av dem som fortplantar sig enbart på könlös väg.
Nackdelarna med könlig förökning
Det må synas förvånande att könlig (sexuell) förökning är så vida spridd som den är. Den är nämligen energimässigt mycket kostsammare än könlös (asexuell) förökning. Vid könlös förökning producerar alla individer avkomma. Vid könlig förökning behövs däremot hos djur vanligen både hanindivider och honindivider. Men det är i regel bara hälften av individerna, nämligen honindividerna, som producerar avkomma. Hanindividerna befruktar bara honindividerna. Ett undantag är hermafroditiska djur, hos vilka samma individ har både hanliga och honliga könsorgan. De flesta hermafroditer självbefruktar sig inte, utan korsbefruktar varandra. Hos dem producerar således alla individer avkomma. Vid könlig förökning hos djur med inre befruktning måste hanar och honor hitta varandra och i regel utföra parningslekar för att kunna föröka sig. Detsamma gäller korsbefruktande hermafroditer. Detta kräver också energi. Djur med yttre befruktning måste synkronisera fortplantningen och producera enorma mängder könsceller, något som är mycket energikrävande.

Fördelarna med könlös förökning
Hos djur sker könlös förökning bland annat genom att delar av ett djur knoppar av nya individer eller genom partenogenes ("jungfrufödsel"). Sådan förökning ger upphov till genetiskt identiska individer som tillsammans utgör en klon. Vid könlös förökning producerar alla individer avkomma. Det behövs inga hanar. Självbefruktande hermafroditism är jämförbar med könlös förökning, eftersom den innebär en kontinuerlig inavel och med tiden ger upphov till genetiskt identiska individer. Eftersom befruktning inte behövs kan djur med könlös förökning ofta föröka sig mycket snabbt. Om det sker en snabb miljöförändring som öppnar nya möjligheter för dem, kan detta vara en fördel. Men på längre sikt är det i regel en nackdel. Varför?
De stora fördelarna med könlig förökning
Vid könlig förökning sker en omkombination av gener dels genom att kromosomer från fadern och modern fördelas på olika sätt till olika individer inom avkomman, dels genom att det under den så kallade meiosen (reduktionsdelningen) sker ett utbyte av gener mellan de båda likadana kromosomerna i ett kromosompar inom den så kallade diploida kromosomuppsättningen. Allt detta gör att individer med helt nya genkombinationer ständigt uppkommer. Detta innebär att gynnsamma mutationer snabbare kan etableras i populationen och att ogynnsamma mutationer lättare kan sorteras bort.
Utan sexuell förökning blir det problem, om en ny gynnsam mutation uppkommer i en individ, som även bär på ogynnsamma mutationer. Om de ogynnsamma effekterna överväger, kommer en sådan individ att få en liten avkomma och sorteras därför bort av det naturliga urvalet. Om de gynnsamma effekterna överväger, kommer de ogynnsamma mutationerna föras vidare som en barlast till följande generationer.
Vid sexuell förökning kan däremot "agnarna sorteras bort från vetet". Då kan en nyuppkommen gynnsam mutation kombineras med andra gynnsamma mutationer, samtidigt som ogynnsamma mutationer sorteras bort. Individer, som bär på de gynnsamma mutationerna och saknar de ogynnsamma, kommer att få en större avkomma än andra. Dessa individer gynnas starkt av det naturliga urvalet. Dessutom finns det en stor genetisk variabilitet hos populationens individer. Detta är en stor fördel, om populationen drabbas av stora miljöförändringar, nya predatorer, parasiter eller sjukdomar. Variabiliteten ger då ett utgångsmaterial för det naturliga urvalet att verka på. Då ökar chansen att en stor andel individer inom en population överlever, genetiskt anpassade till att klara av de nya påfrestningarna.
Sammanfattningsvis skulle vi kunna säga att könlig förökning, med hjälp av det naturliga urvalet, bibehåller gynnsamma mutationer och sorterar bort ogynnsamma. Desutom leder den till en större genetisk variabilitet i populationen, något som kan vara en fördel vid miljöbetingad stress.

Ovanstående teori grundades först på matematiska modeller, men saknade länge experimentellt stöd. Nu har det emellertid kommit flera studier av experimentell evolution, som ger den stöd. En utmaning för teorin är dock fortfarande hjuldjuren inom ordningen Bdelloidea, som tros ha förökat sig könlöst genom partenogenes i minst 80 miljoner år och i dag består av cirka 400 arter.
Sexuellt urval
En annan fördel med könlig förökning är att den möjliggör så kallat sexuellt urval (sexuell selektion) vid partnervalet. Sexuellt urval förkommer dock inte inom alla djurgrupper och kan därför inte vara den viktigaste fördelen med könlig förökning.
Vid den ena typen av sexuellt urval kämpar hanarna om honorna. Det gör de ofta genom att slåss med rivaler med hjälp av horn, näbbar, tänder, klor eller andra hjälpmedel. Ofta räcker det dock med att den starkaste hanen på något sätt demonstrerar sin överlägsenhet. I båda fallen får vinnaren para sig med honan eller honorna.
Vid den andra typen av sexuellt urval väljer honorna sin partner. Hanarna måste då demonstrera sin lämplighet, till exempel genom att sjunga, genom rituell dans, genom att exponera tilldragande färger eller uppseendeväckande utskott på kroppen, genom att ge gåvor eller genom att tillverka ett välbyggt bo. De skickligaste hanarna väljs ut av honorna och får para sig med dem. Hos vissa arter är det hanarna som väljer ut honorna, men detta är inte särskilt vanligt.
Båda selektionstyperna leder till att honorna parar sig med de friskaste hanarna, som är utrustade med de bästa generna, något som i sin tur leder till en mera livskraftig avkomma. Detta har bekräftats genom experiment. Det kan dock i många fall förekomma en konflikt mellan naturligt urval och sexuellt urval. Bjärt färgade hanar upptäcks lättare av predatorer. Hanar med ofunktionella utskott på kroppen, till exempel påfågelhannens extremt långa stjärt, har svårare att fly undan predatorer.
Andra hypoteser. The Red Queen hypothesis
Det finns konkurrerande hypoteser, som försöker förklara fördelarna med könlig förökning på andra sätt, men de måste nu anses spela en mindre roll. De i dessa hypoteser föreslagna mekanismerna skulle dock, för hundratals miljoner år sedan, kunna ha bidragit till uppkomsten av könlig förökning genom att ge vissa fördelar. De skulle också fortfarande i viss mån kunna bidra till att könlig förökning gynnas av det naturliga urvalet. En av dessa hypoteser, "the Red Queen hypothesis", hävdar att den stora genetiska variationen i en population vid könlig förökning skyddar djuren mot parasiter. I en slags evolutionär kapplöpning uppkommer först mutationer hos parasiterna, som gör dem farligare. Detta motverkas av att nya mutationer snabbt kan etableras i värddjurspopulationen och försvara värddjuret mot den nya faran. Enligt denna hypotes upprepas detta ständigt i ett ställningskrig mellan värd och parasit. Termen "the Red Queen hypothesis" syftar på detta ställningskrig. I Lewis Carrolls bok "Through the Looking-Glass" ("Alice i Spegellandet", uppföljaren till "Alice i Underlandet") säger den Röda drottningen: "Now, here, you see, it takes all the running you can do, to keep in the same place."
Fördjupning
Hur påverkas hjärnans kemi hos förälskade? Kärlekens berusning och kval i poesin och biologin. Artikeln finns under rubriken Förnuft, känslor och beteenden – Hur funkar hjärnan, nervsystemet, sinnen och hormoner?
Referenser
- N. Colegrave: Sex releases the speed limit on evolution (Nature 420:664-666, 2002).
- D. Fontaneto et al.: Independently evolving species in asexual bdelloid rotifers (PLoS Biol 5(4): e87).
- M. R. Goddard: Molecular evolution: sex accelerates adaptation (Nature 531:176-177, 2016).
- M. R. Goddard, H. C. J. Godfray, och A. Burt: Sex increases the efficacy of natural selection in experimental yeast populations (Nature 434:636-640, 2005).
- L. Gross: Who needs sex (or nales) anyway? (PLoS Biol 5(4): e99).
- M. J. McDonald, D. P. Rice, och M. M. Desai: Sex speeds adaptation by altering the dynamics of molecular evolution (Nature 531:233-236, 2016).
- L. T. Morran et al.: Running with the Red Queen: host-parasite coevolution selects for biparental sex (Science 333:216-218, 2011).

Vilka djur blir äldst? Och hur gamla kan de bli? Hur gamla kan växter bli? Finns det organismer som inte åldras och i princip skulle kunna leva för evigt? Denna artikel handlar om åldersrekord.
Jag tar här upp de äldsta kända individerna av djur och andra organismer. Detta ger en viss uppfattning om deras maximalt möjliga ålder, det vill säga den ålder de kan nå, om de inte dukar under för rovdjur, svält, parasiter eller sjukdomar. I naturen blir naturligtvis djurens medelålder betydligt lägre än den maximalt möjliga åldern. Ett problem är att vi har mycket lite data om djurens maximala ålder. Ofta handlar det om livslängden hos djur som levt i djurparker, inte vilda djur. Siffrorna nedan blir därmed ofta mycket osäkra. Och vi kommer med stor sannolikhet i framtiden hitta äldre arter och äldre individer av de arter som tas upp här. Artlistorna som förtecknar djuren efter maximal känd ålder kommer således att revideras i framtiden. Notera att jag bara tar upp åldrar på individer vars ålder bekräftats med acceptabel eller hög säkerhet. Det finns en uppsjö av uppgifter om mycket gamla djur som inte kan bekräftas eller som är ren lögn. Inte minst gäller detta om mänskliga åldringar.

De äldsta däggdjuren
Om vi börjar med däggdjur, så är människan ett av de mest långlivade däggdjuren. Hennes maximal möjliga livslängd tros vara cirka 115–120 år. Madame Jeanne Louise Calment (1875–1997) blev 122 år gammal. Hon är den äldsta kända människan och människan den näst äldsta kända däggdjursarten. Det finns ett antal andra uppgifter om människor, som anses ha blivit cirka 120 år gamla. En del av dessa skulle kunna vara korrekta, men bevisen är alltför svaga. Klart är att antalet personer, som blivit äldre än hundra år, har ökat i många länder under de senaste decennierna, förmodligen på grund av bättre levnadsförhållanden och förbättrade medicinska behandlingsmetoder. I Sverige byts numera bindestrecket i personnumret ut mot ett plustecken på hundraårsdagen. Därmed slipper de aktningsvärda åldringarna få adresserad reklam för babyblöjor och annat som ett spädbarn behöver.

Madame Calment hade förmodligen en exceptionell genuppsättning, som möjliggjorde hennes långa liv. Hon uppges ha varit mycket okänslig för stress. Men hon ansträngde sig inte för att leva ett särskilt sunt liv. Hon påstod att hon åt cirka 1 kg choklad i veckan. Hon slutade dock att röka vid 119 års ålder, förmodligen för att hon var så blind att hon inte kunde tända cigaretterna.
Madame Calment sålde år 1965 vid 90 års ålder sin lägenhet i Arles till en advokat. Villkoren var att hon skulle få bo kvar fram till sin död med en årlig livränta på 2 500 franc, varefter advokaten skulle överta lägenheten. Han trodde förmodligen att han snart skulle få flytta in i en ny bostad. Han avled emellertid 1995, 30 år senare, då Madame Calment fortfarande var i livet. Han och hans efterkommande fick betala ett mycket högt pris för bostaden. Så kan det gå för den som försöker lura gamla damer. Men Madame Calment lär bara ha fällt den torra kommentaren: "Man gör ibland dåliga affärer under sitt liv".
Det äldsta med acceptabel säkerhet åldersbestämda däggdjuret är emellertid en grönlandsval (Balaena mysticetus). Personer har i grönlandsvalar hittat harpunblad av sten och elfenben. Inuiterna slutade att använda sådana harpuner i mitten av 1800-talet. Dessa valar kan ha varit åtskilligt mer än 100 år gamla. Harpunfynden föranledde forskare att försöka åldersbestämma grönlandsvalar med hjälp av åldersbetingade förändringar i de proteiner som bygger upp ögats lins. De flesta proteiner i kroppen omsätts ständigt så att de gamla bryts ned och nya bildas. Ett undantag är just proteinerna i ögats lins. Proteinerna längst in i linsen är således kvar sedan linsen anlades i fostret. När forskare undersökte ögonlinserna hos ett antal grönlandsvalar som dödats mellan 1978 och 1996 hittade de fyra valar som bör ha varit äldre än 100 år när de dog. En av dessa valar åldersbestämdes till cirka 211 år. Den valen får betraktas som det äldsta kända däggdjuret.

Människoaporna blir inte lika gamla som människor. För den äldsta med någorlunda säkerhet åldersbestämda gorillan (Gorilla sp.) anges åldern 55,4 år, för orangutangen (Pongo sp.) 59 år, för den äldsta bonobon (Pan paniscus) 5 år och för den äldsta schimpansen (Pan troglodytes) 59,4 år. Gorillorna och orangutangerna är numera uppdelade i vardera två arter. Det framgår inte i källan vilka arter det är. Fysiologiska studier tyder på att schimpanser börjar åldras betydligt tidigare än människor. Det cirkulerar många osannolika uppgifter om schimpansers ålder. Mest apart (sic!) är påståendet att artens mest framstående filmstjärna, Tarzans kompis Cheeta, skulle ha dött 2011 80 år gammal, därmed överlevande sina medspelare Johnny Weissmuller och Maureen O'Sullivan med åtskilliga år.
I allmänhet blir däggdjur äldre ju högre deras kroppsvikt är. Stora däggdjur lever alltså längre än små. Men det finns flera undantag från denna regel. Människan är ett flagrant undantag, eftersom hon blir anmärkningsvärt gammal i förhållande till sin låga kroppsvikt. Det kan finnas evolutionära fördelar med detta. En fördel med ett långt liv för oss människor är att det ges tid att överföra det kulturella arvet från generation till generation. Därmed hinner varje generation med att lära nästa generation en mängd saker, som ökar överlevnadsförmågan. Det behövs, med andra ord, ordentlig utbildning för att lära sig tillverka stenyxor, tända eld, samla ätliga rötter och skjuta med pilbåge. En annan möjlig förklaring till människans långa liv är att hon löper mindre risk att dödas av rovdjur, tack vare sin kroppstorlek, sitt välutvecklade sociala liv och sin avancerade kultur. Då kan det vara en evolutionär fördel att fortplanta sig under flera år och att producera en liten avkomma som vårdas under lång tid.

Nederst ses en dugong (Dugong dugon) som jämte manaterna tillhör ordningen sirendjur (Sirenia). Sirendjuren är, jämte valarna, de enda däggdjur som lever hela sitt liv i vatten, utan att gå upp på land för att fortplanta sig. Dugongen är en växtätare som finns i kustnära tropiska havsområden. Den uppges väga cirka 200–400 kg och kan bli mycket gammal. Den äldsta kända dugongen uppges ha blivit 73 år gammal. Ett utdött sirendjur, Stellers sjöko, levde i norra delen av Stilla Havet, men utrotades på 1700-talet. Den lär ha nått en längd på cirka 8 meter och en vikt på cirka 10 ton. Sjökorna är inte nära släkt med sälarna och inte med valarna heller. Till deras nära släktingar hör faktiskt elefanterna.
Honor av dugonger och manater har på framkroppen ett par spenar, som är formade som kvinnobröst. Det påstås att detta har gett upphov till myten om sjöjungfrur. Sjömän har vid åsynen av djuren trott att de sett varelser med kvinnoöverkropp och fiskstjärt. Efter att ha sett bilden ovan, undrar jag hur pass nyktra dessa män var.
Ordningens namn, Sirenia, syftar på en annan myt. Sirenerna var i grekisk mytologi undersköna kvinnovarelser, som med sin förledande sång lockade sjömän att segla så nära en klippig kust att deras skepp gick i kvav. Den mångförslagne Odysseus undvek faran genom att låta sina sjömän plugga igen öronen med bivax. Själv ville han höra sirensången och lät därför sina män surra fast honom vid masten. Han befallde dem att inte släppa honom fri, förrän sirenerna var utom hörhåll, hur mycket han än bönade och bad.
På listan över de hundra äldsta kända däggdjuren är det bara fyra arter som visats kunna bli mer än hundra år gamla: grönlandsvalen och människan, som diskuterats ovan, samt sillvalen (Balaenoptera physalus; 114 år) och blåvalen (Balaenoptera musculus; 110 år). Som väntat domineras listan av stora djur med hög kroppsvikt. Inte mindre än 27 arter är valar, däribland nio av de tio översta på listan. Den tionde är människan, på andra plats. De fem tyngsta landdäggdjuren återfinns också på listan: afrikansk elefant (Loxodonta sp.; oklart vilken av de två arterna som avses) 65 år; indisk elefant (Elephas maximus) 65,5 år; flodhäst (Hippopotamus amphibius) 61,2 år; trubbnoshörning (Ceratotherium simum) 45 år; spetsnoshörning (Diceros bicornis) 49 år.
Anmärkningsvärt är att 36 av arterna på listan är apor, förutom människan och de ovan nämnda fyra människoaporna, tio gibbonarter samt 22 andra arter från både Gamla och Nya världen, flera av de senare arterna ganska små. De svanslösa gibbonerna tillhör samma överfamilj (Hominoidea) som människor och människoapor. De kan också bli nästan lika gamla som människoaporna (40–60 år). Du bör dock notera den ovan använda listan inte ger ett rättvisande urval av däggdjursarter. Det är svårt att få åldersuppgifter för många arter. Dessutom torde arter som hålls i djurparker vara överrepresenterade. Det är dock klarlagt att inte bara människan, utan också de andra arterna inom ordningen Primates (människoaporna, aporna, spökdjuren och halvaporna) tenderar att leva längre, än vad vi skulle förvänta oss utifrån deras kroppsvikt. De kan i och för sig ha en ganska hög basal energiomsättning i vila. Men i en studie har forskarna funnit att de har en jämförelsevis låg total energiomsättning under loppet av ett dygn. Detta skulle kunna leda till en långsam livscykel, inkluderande tillväxt, fortplantning och åldrande.

Mest anmärkningsvärt är att forskarna hittar tre fladdermusarter bland de hundra äldsta kända däggdjuren, äldst av dem ett exemplar av Brandts fladdermus (Myotis brandti), som blev 41 år gammalt. Fladdermöss blir exceptionellt gamla i förhållande till sin mycket låga kroppsvikt. Orsaken är inte helt klarlagd, men enligt en hypotes har det att göra med att de måste omsätta enormt mycket energi vid flygning och därmed också förbruka väldigt mycket syre. Syre är, i synnerhet i höga doser, giftigt genom att det ger upphov till så kallade fria radikaler, som skadar cellernas molekyler. Vi skulle därför förvänta oss att höga halter av fria radikaler förkortade fladdermössens livslängd. Forskarna har påvisat genmutationer, som ger dem en ovanligt bra förmåga att reparera sitt DNA. De tycks också vara extra bra på att oskadliggöra fria radikaler. Möjligen har fladdermössen varit tvungna att utveckla ett extra bra försvar mot fria radikaler för att leva länge. Men det finns en rad studier som pekar på att fria radikaler inte påskyndar åldrande. Numera har de flesta forskare övergivit teorin att de förkortar livslängden.
Fladdermössen har också ett immunsystem, som gör att de inte drabbas av skadliga inflammationer. Vid dödliga virusinfektioner är det oftast inte viruset i sig som skadar kroppen utan ett alltför kraftigt inflammatoriskt immunförsvar. Fladdermöss härbärgerar många virus, som inte skadar dem, men som blir enormt farliga, när de överförs till människor. Ett känt exempel är ebolaviruset.
De äldsta fåglarna
Däggdjur lever vanligen längre, ju större de är. Denna tendens finns också hos fåglar. En fågel tenderar däremot att leva längre än ett lika stort däggdjur. Men det finns flera undantag. Notera att vi här, precis som ovan, talar om den maximalt möjliga livslängden och att det ofta rör sig om fåglar i fångenskap. I naturen är livet hårt, vilket gör att djur där nästan aldrig når sin maximalt möjliga livslängd.
Det är intressant att de flygande ryggradsdjuren, fladdermöss och fåglar, båda utmärker sig för att nå en hög ålder. I vila har gruppen fladdermöss en högre energiomsättning per gram kroppsvikt, än vad vad skulle förvänta oss av däggdjur med deras kroppsvikt. Fåglarna som grupp har högre energiomsättning per gram kroppsvikt än däggdjuren. Det innebär att en fågel nästa alltid har en högre energiomsättning än ett däggdjur med samma kroppsvikt. Både fåglar och fladdermöss har en extremt ökad energiomsättning när de flyger. Som nämnts ovan leder en hög energiomsättning och därmed en hög syreförbrukning till att det bildas avsevärt fler skadliga fria radikaler. Vi skulle förvänta oss att detta ledde till skador som förkortade de flygande djurens maximalt möjliga ålder. I stället är det tvärtom, dessa djur lever mycket länge. Forskarna har också spekulerat att flygande djur lättare flyr från rovdjur och att det naturliga urvalet hos dem därför skulle gynna en högre medellivslängd, under vilken de kan fortsätta att reproducera sig.
De högsta med acceptabel säkerhet belagda åldrarna för fåglar över huvud taget är 81 år för en kakaduapapegoja (arten Cacatua leadbeateri), 79 år för en andinsk kondor (Vultur gryphus), 70 år för en knölsvan (Cygnus olor), 70 år för en näshornsfågel (Bucorvus leadbeateri) och 69 år för en korp (Corvus corax). Alla dessa fåglar levde i fångenskap.

Här är några med acceptabel säkerhet belagda åldersrekord för några andra fåglar, en del för djur i naturen och en del för djur i fångenskap: kungsalbatross (Diomedea epomophora) 58 år; grå stormfågel (Fulmarus glacialis) 51 år; afrikansk struts (Struthio camelus) 50 år; jakopapegoja (Psittacus erithacus) 49,7 år; gråtrut (Larus argentatus) 49 år; kungsörn (Aquila chrysaetos) 48 år; bläsgås (Anser albifrons) 47 år; flamingon Phoenicopterus roseus 44 år; vit stork (Ciconia ciconia) 39 år; sillgrissla (Uria aalge) 42,8 år; bofink (Fringilla coelebs) 29 år; bredstjärtad kolibri (Selasphorus platycercus) 14 år; lövsångare (Phylloscopus trochilus) 11,8 år. För flera av dessa fåglar finns högre åldrar angivna i källor som inte kan anses som tillräckligt trovärdiga.
Det är huvudsakligen stora fåglar som når hög ålder, men tendensen är inte särskilt tydlig. Till de grupper som kan bli mycket gamla hör papegojor, rovfåglar, näshornsfåglar och albatrosser. De flesta av dem är stora fåglar. Flera stora fåglar kan bli lika gamla som elefanter. På listan över de hundra äldsta kända fåglarna är 25 arter papegojor och 19 arter stora rovfåglar (framför allt örnar, gamla världens gamar och nya världens gamar). Det stora antalet papegojor kan dock delvis bero att vi har många uppgifter, eftersom så många av dem hålls som burfåglar.
Kråkfåglarna och papegojorna är kända för en högt utvecklad kognitiv förmåga ("intelligens"). Med undantag av korpen tycks kråkfåglar dock inte bli lika gamla som papegojor. De högsta med acceptabel säkerhet belagda rekorden är för skatan (Pica pica) 21,7 år, för kråkan 19,2 år och för korpen 69 år.
Men flera små fåglar kan också nå en respektingivande ålder, notera bofinken, trädgårdssångaren och stormfågeln i listan ovan. Notera också kolibrin, som kan bli 3-4 gånger äldre än ett däggdjur med motsvarande kroppsvikt.
De äldsta kräldjuren
Det finns stora landsköldpaddor som kan bli mycket gamla. Det finns ett trovärdigt belägg för en ålder på minst 177 år för Geochelone nigra (numera uppdelad i flera arter inom slåktet Chelonoidis) från Galapagos och ett på minst 152 år för Geochelone gigantea från Seychellerna. Dessa två individer är de äldsta kända kräldjuren. Några anekdotiska uppgifter anger att vissa jättesköldpaddor kan bli mer än 200 år gamla, men de måste betraktas som obestyrkta, om än sannolika. Den mest berömda av alla sköldpaddor, Lonesome George på Galapagosöarna, blev cirka 100 år gammal. Han bar sitt dystra namn därför att han var den siste av sin art.

På listan över de hundra äldsta kända reptilerna dominerar sköldpaddorna stort, möjligen på grund av deras låga energiomsättning. Av listans arter är 52 sköldpaddor, både stora och små arter. De fem arter, som uppges nå en ålder över 100 år, är alla sköldpaddor. Det äldsta belagda krokodildjuret uppges vara en mississippialligator på 73,1 år. Den äldsta ormen uppges vara en kungspyton på 47,5 år. Det har påståtts att komodovaraner kan bli 62 år, men beläggen är alltför svaga. Därmed kvalar, hör och häpna, en kopparödla (Anguis fragilis; ormslå, "kopparorm") in som den äldsta belagda ödlan med 54 år. Denna ödla är vanlig i Sverige. Intressant nog kommer den nya zeeländska tuataran (bryggödlan) på femte plats med 90 år, efter de fyra mer än hundraåriga sköldpaddorna. Den är ingen ödla utan bildar en alldeles egen reptilordning.
De äldsta groddjuren
Bland groddjuren utmärker sig en art framför alla andra, olmen (Proteus anguinus) med en ålder på 102 år. Beläggen är indirekta, men anses ändå acceptabla. Denna grottlevande salamander genomgår inte, som nästan alla andra groddjur, en metamorfos från larv till vuxen. Larverna blir i stället könsmogna, så kallad neoteni. Forskarna har inte upptäckt tecken till åldrande hos olmen. Den tros vara ett av de djur som skulle kunna leva för evigt, om den inte dödas av yttre påverkan, se längre ner på sidan. Den japanska jättesalamandern (Andrias japonicus) uppges kunna bli 55 år och den afrikanska grävoxgrodan (Pyxicephalus adspersus) 45 år. Dessa tre arter är de äldsta kända groddjuren. I övrigt finns det många stjärtgroddjur och stjärtlösa groddjur som uppges nå en ålder mellan 20 och 40 år.

De äldsta fiskarna
År 2016 publicerades en undersökning av åldern hos håkäringen (Somniosus microcephalus), en haj som lever i Nordatlantens kalla arktiska vatten, på vintern också vid Västkusten. Den växer bara cirka 1 cm om året i längd, men kan ändå bli 5 m lång eller längre. Den rör sig normalt mycket långsamt, utom då den fångar sina byten. Forskare bestämde åldern genom att kol-14-datera proteiner inne i ögats lins, en metod som mäter nedbrytningen av en radioaktiv kolisotop. Linsproteinerna bildas under fosterstadiet och omsätts inte, vilket innebär att kolatomerna i dem är lika gamla som hajen. Den äldsta hajen var 392 ± 120 år gammal. Standardavvikelsen och därmed osäkerheten i kol-14-mätningar är stor, men denna haj är med allra största sannolikhet det äldsta ryggradsdjur, vars ålder bestämts med rimlig säkerhet. Förvånansvärt nog tydde data på att håkäringar blir könsmogna först vid cirka 150 års ålder. En sannolik orsak till dessa hajar lever så länge är att de har en mycket låg energiomsättning i det kalla vattnet. Möjligen kan det också vara så att kylan aktiverar vissa gener som ökar livslängden. Effekten av sådana gener har påvisats hos rundmasken Caenorhabditis elegans och de finns sannolikt hos många andra djur. Den efter håkäringen äldsta med acceptabel säkerhet belagda broskfisken (Chondrichthyes; hajar, rockor och helhuvudfiskar) är konstigt nog med 75 år den lilla pigghajen (Squalus acanthias), som finns vid Västkusten. För den största hajen, valhajen (Rhincodon typus), uppges bara 54 år och för vithajen (Carcharodon carcharias) 50 år. Det beror antagligen på brist på data.

Förutom håkäringen, finns det ett tjugotal andra fiskar som uppges kunna bli 100 år gamla eller äldre. Som nummer två och tre på listan efter håkäringen kommer kungsfiskarterna Sebastes aleutianus med 205 år och Sebastes borealis med 157 år. Som nummer fyra kommer störarten Acipenser fulvescens med 152 år. Synnerligen märkligt är att inte mindre än 34 arter på listan över de hundra äldsta kända strålfeniga fiskarna (Actinopterygii) tillhör kungsfisksläktet Sebastes, däribland med 40 år mindre kungsfisk (Sebastes viviparus), som finns vid Västkusten. Släktet Sebastes åldras kanske inte. Märkligt är också att nio av de 24 arterna inom familjen störfiskar (Acipenseridae) finns med på listan, flera av dem högt upp. De strålfeniga fiskarna, i synnerhet gruppen Teleostei, är den utan jämförelse artrikaste nutida fiskgruppen. För tofsstjärtfisken Latimeria chalumnae (kvastfeningen, det berömda "levande fossilet") uppges en maximal känd ålder på 48 år. För rundmunnar och lungfiskar hittar jag inga uppgivna åldrar över 20 år.
En anledning till att flera kräldjur och fiskar kan bli mycket gamla är kanske att de ofta växer hela livet, om än långsammare ju äldre de blir. Många av dem kan dessutom fortplanta sig vid hög ålder.
Ryggradslösa djur och djur med evigt liv
De stora sköldpaddorna, grönlandsvalen och håkäringen är de äldsta åldersbestämda ryggradsdjuren. Men det kan ju finnas andra åldringar som vi ännu inte upptäckt. Den äldsta åldersbestämda djurindivid vi känner till är en islandsmussla (Arctica islandica) som var 507 år gammal. Det finns en uppgift om att långsamt växande svampdjur (arten Scolymastra joubini) i havet kring Antarktis är 15 000 år gamla, men detta har ifrågasatts. Uppgiften att individer av svampdjuret Monorhaphis chuni i Kinesiska sjön skulle vara cirka 11 000 år gamla kan också ifrågasättas. Det är emellertid möjligt att dessa svampdjur kan bli betydligt äldre än den ovan nämnda musslan, kanske 1000-tals år gamla. Det kan vara så att de inte åldras alls.
De äldsta åldersbestämda djuren skulle emellertid kunna vara rundmaskar (nematoder). Dessa maskar har dock, till skillnad från islandsmusslan ovan, tillbringat nästan hela sitt liv frusna till is, i ett vilstadium som kallas kryptobios. Ryska forskare har hittat dem djupt nere i den sibiriska permafrosten i lager, som med kol-14-metoden åldersbestämts till cirka 32 000 och cirka 42 000 år. Maskarna "vaknade" och fortplantade sig efter att ha tinats och förvarats i rumstemperatur. De låg så djupt att de aldrig tidigare tinats. En rad säkerhetsåtgärder vidtogs för att undvika kontamination med nu levande maskar. Fynden är dock så extraordinära att vi skulle vilja ha dem bekräftade i en ny undersökning på ett annat labb.

En del menar att vissa djur inte åldras och teoretiskt skulle kunna leva hur länge som helst. Detta skulle kunna gälla vissa reptiler och fiskar samt flera ryggradslösa djur. För vissa arter finns det studier som visar att de har försumbart åldrande. Till dem hör en dossköldpaddeart (Terrapene carolina), två andra sköldpaddsarter, olmen (Proteus anguinus), en kungsfiskart (Sebastes aleutianus), en sjöborrart (Strongylocentrotus franciscanus) och den ovan diskuterade islandsmusslan (Arctica islandica). Om uppgifterna stämmer torde dessa djur vara utrustade med mycket välutvecklade försvarsmekanismer mot de processer som gör att djur åldras.

Nässeldjuret Turritopsis nutricula (klassen Hydrozoa), kan genom så kallad transdifferentiering omvandlas från en frilevande medusa till en fastsittande polyp. Polypen bildar nya medusor. Den normala utvecklingen hos nässeldjur är den motsatta. Ur ägget kläcks en larv som omvandlas till en polyp. Polypen bildar medusor genom avknoppning. Medusorna dör efter att ha förökat sig på könlig väg. Vi skulle kunna säga att Turritopsis lever ett evigt liv: först som polyp, sedan som medusa och sedan som polyp igen, i ett ständigt kretslopp. Den dör inte, om den inte dukar under för predatorer eller sjukdomar. Men eftersom en polyp knoppar av flera medusor förökar sig djuret genom att ge upphov till nya medusor, som alla har potentialen att leva ett evigt liv.

Växter, svampar och andra organismer
Hur är det då med andra organismer än djuren? Det brukar anges att den äldsta kända individen av en organism är ett exemplar av tallarten Pinus longaeva som år 2012 var 5 062 år gammalt. Denna tallart växer i Klippiga bergen i USA. Ålder på träd kan vi ju bestämma mycket noga genom att räkna årsringarna i stammen. I Dalarna har forskare hittat granar med buskartat växtsätt som är mycket gamla. Den äldsta granen beräknas vara 9 550 år gammal. Åldern har bestämts genom kol-14-datering av döda vedrester i marken under granen. De nu levande stammarna, individerna, är betydligt yngre. Eftersom granen förökat sig genom rotslående grenar, varvid nya stammar bildats, är den en klon, inte en individ, se nästa stycke. Gränsen mellan individ och klon kan dock vara flytande när det gäller växter. Granfynden visar i varje fall att granen efter istiden invandrade till Sverige ca 8 000 år tidigare än vi tidigare trodde och att den kan ha kommit från väster i stället för från öster.

Hos växter och djur som förmerar sig vegetativt (med könlös förökning) tillhör alla individer samma klon och är genetiskt i det närmaste identiska. Det har hävdats att en klon är en enda individ som i princip kan leva hur länge som helst. Men begreppet individ förlorar då helt sin mening. Vi kan inte räkna en klon som en individ. Det finns dock flera exempel på kloner av växter som beräknats vara tusentals år gamla. Det handlar då om klungor av plantor eller träd. Ett känt exempel är en aspklon, som finns i Utah i USA. Den anses bestå av mer än 47 000 individer (det vill säga stammar), beräknas väga mer än 6 miljoner kg och anses vara uppemot 10 000, kanske till och med 1 miljon, år gammal.

Ett mycelium av en honungskivlingsart (släktet Armillaria) i USA beräknades vara mer än 1 500 år gammalt, väga mer än 10 000 kg och täcka mer än 15 hektar. Molekylärbiologisk provtagning visade att mycelets celler var genetiskt identiska och mycelet betraktades som en individ. Senare har forskare hittat ännu större honungsskivlingsmycel. Men det är oklart om ett stort svampmycel en individ eller en klon. Potatis förökas vegetativt genom att vi sätter ner knölarna i jorden. Varje potatissort har, åtminstone teoretiskt, chansen att leva ett evigt liv. På ett liknande sätt förhåller det sig med bakterier och andra encelliga organismer. De åldras inte, utan de delar sig ständigt, ger då upphov till nya celler och kan därför i princip sägas leva i evighet. Åldrande är alltså något som bara kan finnas hos flercelliga organismer med könlig förökning. Hos vissa bakterier och hos jästsvampar har forskare dock hittat förändringar som tolkats som åldrande.
Flera miljoner år gamla bakterier?
Forskare har hitta levande bakterier mycket djupt ner i marina sediment och sedimentära bergarter. Sådana mikroorganismer utgör kanske en mycket stor del av Jordens biomassa. Dessa bakterier lever på sparlåga med en oerhört låg ämnesomsättning. Det är oklart om de förökar sig genom delning. Om de delar sig gör de det kanske med hundratals års mellanrum eller mer. De skulle också kunna befinna sig i ett vilstadium utan att dela sig. I så fall återspeglar deras ämnesomsättning bara de processser som behövs för att hålla dem vid liv. De har hittas i bergarter som är miljontals år gamla. Mer forskning behövs inom detta område. Men dessa bakterier skulle kunna vara de äldsta kända levande varelserna.
In memoriam: Lonesome George
Sommaren 2012 inflöt det dystra meddelandet att Lonesome George avlidit, sörjd av alla som värnar biodiversiteten på vår jord. Ecuadors president hedrade hans minne i ett tal till nationen. Vid obduktionen kunde zoologer inte utröna dödsorsaken. George var förmodligen cirka 100 år gammal när han dog. Försök har gjorts att föra Georges gener vidare genom att låta honom fortplanta sig med honor av besläktade arter av galapagossköldpaddor. Försöken har emellertid misslyckats. Det sista exemplaret av pintasköldpaddan (Chelonoidis abingdoni) är därmed borta. Zoologer har dock sparat frusna cellprover och vävnadsprover. Möjligen kan arten i en framtid återupplivas med hjälp av stamceller från dessa prover.
Pintasköldpaddornas förfäder var förmodligen individer av arten Chelonoidis hoodensis som för cirka 300 000 år sedan emigrerade från ön Española. Väl på ön Pinta genomgick dessa sköldpaddor en evolutionär anpassning till den nya miljön. Samma sak gäller de tio kvarvarande skölddpaddsarterna på Galapagosöarna. De har alla anpassat sig till miljön på sin ö, till exempel till de växter som utgör deras föda. Detta har bland annat lett till att olika arter har olika form på ryggskölden. Skillnaderna iakttogs redan av Charles Darwin som besökte Galapagos under sin resa med fartyget Beagle. Detta var en av de iakttagelser som inspirerade honom till att senare presentera utvecklingsläran, en av de mest fruktbärande vetenskapliga teorierna någonsin.
En arts utdöende kan leda till allvarliga effekter på ett ekosystem och vara en förlust för både grundforskning och tillämpad forskning. Men det är också en kulturell förlust, precis som förlusten av ett språk eller en bok. Byt ut "man" mot "species" (sv. "art") i den engelske poeten John Donnes diktrader: "No man is an island, / Entire of itself. / Each is a piece of the continent, / A part of the main".
Fördjupning
- Varför är insekter så små? Varför är däggdjur och fåglar stora? Du hittar artikeln under rubriken Varför är vissa djur stora medan andra är små?.
- Hur stor kan du vara och. ändå flyga? Artikeln finns under rubriken Varför är vissa djur stora medan andra är små?.
- Hur nya ben och svansar ersätter gamla. Regeneration: återväxt av amputerade kroppsdelar. Du läser artikeln under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?.
- Världens största ödla, komodovaranden. Läs artikeln under rubriken Märkliga och farliga djur.
- Den biologiska klockan, tuatarans tredje öga och filosofen Descartes. Under rubriken Förnuft, känslor och beteenden – Hur funkar hjärnan, nervsystemet, sinnen och hormoner? kan du läsa artikeln.
Referenser
- AnAge: The Animal Ageing and Longevity Database (Human ageing genomics resources, hämtad 29 augusti 2016).
- Anonymous: A 120-year lease on life outlasts apartment heir (The New York Times, 29 december 1995).
- P. G. Butler et al.: Variability of marine climate on the north Icelandic shelf in a 1357-year proxy archive based on growth increments in the bivalve Arctica islandica (Palaeogeography, Palaeoclimatology, Palaeoecology 373:141-151, 2013).
- E. C. Charnov och D. Berrigan: Why do female primates have such long lifespans and so few babies? or Life in the slow lane (Evolutionary Anthropology 1:191-194, 1993).
- P. K. Dayton et al.: Recruitment, growth and mortality of an Antarctic hexactinellid sponge, Anoxycalyx joubini (PLoS ONE 8[2]: e56939, 2013).
- B. E. Flanary och G. Kletetschka: Analysis of telomere length and telomerase activity in tree species of various life-spans, and with age in the bristlecone pine Pinus longaeva (Biogerontology 6:101-111, 2005).
- A. A. Fushan et al.: Gene expression defines natural changes in mammalian lifespan (Aging Cell 14:352-365, 2015).
- J. C. George et al.: Age and growth estimates of bowhead whales (Balaena mysticetus) via aspartic acid racemization (Canadian Journal of Zoology 77:571-580, 1999).
- Gerontology Research Group (hämtad 15 augusti 2016).
- K. P. Jochum et al.: Siliceous deep-sea sponge Monorhaphis chuni: a potential paleoclimate archive in ancient animals (Chemical Geology 300-301:143-151, 2012).
- A. King: Super bats. What doesn't kill them, could make us stronger (New Scientist, 13 februari 2016).
- L. Kullman: Norway spruce present in the Scandes Mountains, Sweden at 8 000 BP: new light on the Holocene tree spread (Global Ecology and Biogeography Letters 5:94-101, 1996).
- R. M. Lanner och K. F. Connor: Does bristlecone pine senesce? (Eperimental Gerontology 36:675-685, 2001).
- Lonesome George (Galapagos Conservancy, hämtad 29 augusti 2016).
- M. Mangel et al.: Evolutionary analysis of life span, competition, and adaptive radiation, motivated by the pacific rockfishes (Sebastes) (Evolution 61:1208-1224, 2007).
- J. B. Mitton och M. C. Grant: Genetic variation and the natural history of quaking aspen (Bioscience 46:25-31, 1996).
- J. Nielsen et al.: Eye lens radiocarbon reveals centuries of longevity in the Greenland shark (Somniosus microcephalus) (Science 353:702-704, 2016).
- S: Piraino et al.: Reversing the life cycle: medusae transforming into polyps and cell transdifferentiation in Turritopsis nutricula (Cnidaria, Hydrozoa) (Biological Bulletin 190:302-312, 1996).
- H. Pontzer et al.: Primate energy expenditure and life history (Proceedings of the National Academy of Sciences 111: 1433-1437, 2014).
- Rocky Mountain Tree Ring Research Group (updated January 2016).
- R.D. Rosen: Lie of the jungle: the truth about Cheeta the chimpanzee (Washington Post, 7 december 2008).
- A. V. Shatilovich et al.: Viable nematodes from late Pleistocene permafrost of the Kolyma River Lowland (Doklady Biological Sciences 480:100-102, 2018).
- M. L. Smith et al.: The fungus Armillaria bulbosa is among the largest and oldest living organisms (Nature 356:428-431, 1992).
- J. R. Speakman: Body size, energy metabolism and lifespan (Journal of Experimental Biology 208:1717-1730, 2005).
- E. J. Stewart et al.: Aging and death in an organism that reproduces by morphologically symmetric division (PLoS Biol 3[2]: e45).
- E. Trembath-Reichert et al.: Methyl-compound use and slow growth characterize microbial life in 2-km-deep subseafloor coal and shale beds (Proceedings of the National Academy of Sciences 114:E9206-E9215, 2017).
Varför måste människor och andra djur förr eller senare dö? Varför lever vissa djur längre än andra? Vi vet mycket lite om vad som orsakar åldrandet. Men det finns många teorier, flera av dem stödda av experiment. Vi kan förmoda att orsakerna är flera. Här följer en allmän diskussion av detta intressanta ämne.
Åldrandet kännetecknas av att kroppens celler börjar fungera sämre. Risken att drabbas av sjukdomar ökar när vi blir äldre, men åldrandet är i sig ingen sjukdom. Forskarna förmodar att åldrande orsakas av det blir fler skador på kroppscellernas stora molekyler ju äldre vi blir och att det är dessa skador som gör att kroppen fungerar sämre och att celler dör. Det kan också vara så att åldrade celler med speciella egenskaper ökar i antal, när vi blir äldre, och gör att kroppens organ fungerar sämre. Hos bland annat möss, bananflugor och rundmasken Caenorhabditis elegans har forskare identifierat flera genvarianter, som ökar livslängden genom att bland annat reglera celldöd och olika typer av stress.
En rundmask ger kunskap om människans åldrande
Vuxna rundmaskar är cirka 1 mm långa. Masken kan gå igenom hela sin livscykel på cirka 3 dygn: från ägg, via fyra larvstadier och till könsmogen vuxen individ. Dess livslängd är i medeltal cirka 2–3 veckor, maximalt cirka 8 veckor. Den tillhör en helt annan gren på djurens släktträd än däggdjuren. Men dess arvsmassa har förvånande stora likheter med människans. Forskarna har kunnat förlänga dess livslängd genom att förändra flera gener och genom att behandla den med biologiskt aktiva ämnen. Flera av dessa fynd anses vara relevanta också för människan.
Caenorhabditis elegans har flera fördelar som försöksdjur. Viktigast är att den alltid har lika många celler av varje celltyp och att forskarna har identifierat alla dessa celler. Alla individer är alltså uppbyggda på exakt samma sätt. Andra fördelar är att masken är lättodlad, har en kort livscykel och är helt genomskinlig under ett mikroskop. Det senare framgår av videon, i vilken flera organ kan urskiljas. Masken saknar hjärt-kärlsystem. Den regelbundet slående strukturen på videon är inte ett hjärta, utan en klaff som avslutar svalget. Maskarna vidgar svalget och suger in bakterier som mals sönder av klaffen, innan de hamnar i tarmen. Det ser ut som om en ljusblixt mot slutet av videon skrämmer maskarna, så att de försvinner ur synfältet.
Kan åldrande förklaras evolutionärt?
Det är faktiskt möjligt att åldrandet är inprogrammerat i arvsmassan. I så fall är åldrandet en utvecklingsprocess, på samma sätt som barnets utveckling till en vuxen människa. Sannolikare är kanske att det naturliga urvalet hos äldre djur inte gynnar upprätthållandet av mekanismer som försvarar cellerna mot skador. Det kan vara djur som är så gamla att de redan reproducerat sig. Det kan också vara djur vars ålder avsevärt överstiger medellivslängden i naturen, där rovdjur, födobrist och sjukdomar gör att de flesta djur dör långt innan de nått den maximalt möjliga livslängden.
Kroppsvikten är den viktigaste av de faktorer som påverkar däggdjurs livslängd. En möjlig förklaring till detta är att stora djur löper mindre risk att dödas av rovdjur, tack vare sin kroppstorlek. Då kan det vara en evolutionär fördel att fortplanta sig under flera år och att producera en liten avkomma som vårdas under lång tid. Det naturliga urvalet skulle med andra ord kunna gynna ett långt liv och ett sent åldrande hos stora däggdjur.
Många kräldjur och fiskar lever också längre än däggdjur av motsvarande kroppsvikt. De kan till och med uppvisa ett "negativt åldrande" i och med att äldre djur har en större förmåga att fortplanta sig. De flesta av dessa djur verkar dock dö så småningom. Några av dem ha dock visat sig ha ett försumbart åldrande. Kanske finns det många fler inte undersökta djur med försumbart åldrande, både ryggradsdjur och ryggradslösa djur.
Under den kenozoiska eran var däggdjuren småväxta och föll förmodligen ofta offer för rovdjur bland dinosaurierna. Kanske var det under denna tid en fördel för dem att snabbt skaffa sig en stor avkomma. Då kunde tillräckligt många ungar överleva och fortplanta sig. Det är samma strategi som gäller för nutida möss och många andra små däggdjur. Men med denna strategi gynnar det naturliga urvalet inte en hög ålder. En spekulation är, att däggdjuren då förlorade mekanismer som motverkar åldrande och senare haft svårt att åter utveckla dem.
Varför kan fåglar, kräldjur och fiskar bli äldre än däggdjur?
Hos däggdjur är det så att större djur vanligen lever längre. En afrikansk elefant kan leva i åtminstone 50–60 år, men en mus lever bara i högst 2–3 år. Med livslängd menar vi här den tid, efter vilken djuren dör av ålderdomssvaghet. I naturen dukar de flesta möss under, till exempel för rovdjur, innan de blivit 2–3 år gamla. Regeln att större djur lever längre har dock flera undantag. Sålunda blir vi människor lyckligtvis mycket äldre än vi skulle förvänta oss med hänsyn till vår relativt låga kroppsvikt. Detsamma gäller fladdermöss. Fåglarna som grupp tenderar att leva längre än däggdjuren. Detta innebär att en fågel vanligen lever längre än ett lika stort däggdjur med samma kroppsvikt. Forskare har spekulerat att flygande djur lättare flyr från rovdjur och att det därför kan vara möjligt för dem att ha en högre medellivslängd under vilken de kan fortsätta att reproducera sig.
Finns det en oöverstiglig övre gräns för människans livslängd?

Före år 1950 är den viktigaste orsaken en kraftigt minskad barnadödlighet, under senare decennier en minskad dödlighet bland äldre. Det senare beror på en minskad dödlighet i framför allt hjärt-kärlsjukdomar, särskilt hjärtinfarkt och stroke, och till en mindre del i cancer. Enligt prognoserna kommer medellivslängden fortsätta att öka, kanske till 90 år eller mer år 2060.
Flera demografiska studier antyder dock att det finns en naturlig övre gräns för människans livslängd, kanske maximalt 115–120 år. Medellivslängden har förvisso ökat under de senaste 150 åren på grund av medicinska framsteg och förbättrade levnadsförhållanden. Men den maximala livslängden har inte ökat under de senaste decennierna. Vi skulle då kunna tro att den maximala livslängden är genetiskt bestämd, precis som fosterutvecklingen. Så behöver det dock inte vara, i stället skulle livslängden kunna begränsas av det naturliga urvalet.
Det finns ingen anledning att anta att det naturliga urvalet skulle gynna en livslängd på mer än 90–100 år. Vi kan jämföra en människa med en bilmotor. Under större delen av livet är det en evolutionär fördel att ha effektiva biologiska reparationsmekanismer, således ordentlig inre service. Vid mycket hög ålder har människor slutat att fortplanta sig och kan inte heller hjälpa barn och barnbarn att överleva och fortplanta sig. Då skulle vi kunna tänka sig att reparationsmekanismerna inte längre gynnas av det naturliga urvalet. Den inre servicen upphör och allla kroppens organ, det vill säga alla motorns delar, fungerar allt sämre och sämre. Till slut upphör motorn helt att fungera.
Det finns emellertid också demografiska studier som tyder på att det inte finns en naturlig övre gräns för människans livslängd. Risken att dö ökar exponentiellt med ökande ålder. Detta innebär att risken att dö nästa år blir större ju äldre du blir. I en studie av italienska åldringar hävdade forskarna emellertid att riskkurvan planar ut efter 105 års ålder. Efter 105 års ålder gammal skulle i så fall risken att dö mellan två födelsedagar vara densamma, hur gammal du än blir, nämligen 50 procent. Detta skulle innebära att människor teoretiskt skulle kunna bli hur gamla som helst. Men med tanke på den höga årliga dödsrisken skulle sannolikheten att bli avsevärt äldre än 105 år vara mycket liten. Frågan om vi har en maximal livslängd eller inte är emellertid definitivt inte avgjord.
Eftersom så många organ sviktar, menar många att det är mycket svårt att öka den maximala livslängden genom medicinska åtgärder. Å andra sidan kan ingen förutsäga den medicinska utvecklingen. Och forskarna har lyckats öka livslängden avsevärt hos försöksdjur med tämligen enkla behandlingar. Åsikterna går isär. Som ovan nämnts, är det fortfarande oklart om det finns en övre gräns för människans livslängd. De flesta menar dock att det sannolikt finns utrymme att förlänga den friska delen av människolivet, då livskvaliteten fortfarande är hög.
Sociala faktorer påverkar medellivslängden. Personer som studerat på högskola eller universitet lever längre än de som bara har grundkoleutbildning. Det finns också studier som tyder på att medellivslängden i rika länder är högre om skillnaderna i inkomst är låga.
Är åldrande kopplat till energiomsättning och fria radikaler?
Det har antagits att det finns ett samband mellan djurens energiomsättning och den ålder de i bästa fall kan uppnå. Med energiomsättning menas här den hastighet med vilken ämnen i födan eller i kroppens förråd bryts ner så att energi i en för kroppen användbar form bildas. En teori är att åldrandet orsakas av så kallade fria radikaler som bildas vid energiomsättningen. Energiomsättningen sker med hjälp av syre vid cellandningen. Syre är en mycket reaktiv molekyl. Det är faktiskt inte bara nyttigt, utan också ett gift som gör att fria radikaler bildas. Fria radikaler är reaktiva ämnen som skadar organismens celler. Enligt den ovan nämnda teorin skulle skador orsakade av fria radikaler leda till åldrande. Dessutom skulle en låg energiomsättning leda till färre skador orsakade av fria radikaler och därmed ett längre liv.
Men de fria radikalernas roll har starkt ifrågasatts. Det finns flera enzymer som skyddar mot fria radikaler. En del studier av försöksdjur har visat att ökad aktivitet av sådana enzymer inte ökar djurens livslängd. Antioxidanter är ämnen som eliminerar fria radikaler. Flera av dem finns i födan. Ökad tillförsel av antioxidanter har inte gett positiva effekter hos människor. Vissa av dem tycks i stället ha ökat dödligheten. Detta gäller bland annat vitamintabletter innehållande β-karoten och vitamin E, när det inte föreligger någon brist på dessa ämnen. I en studie har antioxidanter visat sig motverka de hälsobefrämjande effekterna av motion. Vissa fria radikaler deltar i signalmekanismer inuti celler. Det finns till och med data som tyder på att fria radikaler stimulerar försvaret mot stress. Det finns en rad studier som pekar på att fria radikaler inte påskyndar åldrande. Numera har de flesta forskare övergivit teorin att de förkortar livslängden.
När forskare jämför energiomsättningen hos olika djur, mäter de oftast basalmetabolismen (BMR). Basalmetabolismen är energiomsättningen i kilokalorier per minut hos ett fastande djur i vila vid en behaglig temperatur i omgivningen. En mus har mycket högre basalmetabolism per gram kroppsvikt än en elefant. Detta innebär att 1 000 kg möss måste äta mycket mer än en elefant på 1 000 kg, annars skulle mössen inte kunna upprätthålla sina höga energiomsättningar. Musen blir ju heller inte lika gammal som elefanten. Detta skulle kunna tolkas så att djuren blir äldre, ju lägre energiomsättning de har per gram kroppsvikt. Men fåglar har högre energiomsättning mätt per gram kroppsvikt än däggdjur, och ändå lever de längre. Det är förmodligen den höga kroppsvikten i sig, inte den låga energiomsättningen, som gör att stora djur lever längre än små. Detta skulle kunna bero på att stora djur löper mindre risk att dödas av rovdjur i unga år. Därför kan de ha en långsammare livscykel.
Primaterna är den däggdjursordning som inkluderar halvapor, spökdjur, apor, människoapor och människor. Primaters basalmetabolism, mätt per gram kroppsvikt, är vanligen precis så hög som vi skulle förvänta sig hos ett däggdjur. Ändå tenderar en primat att leva längre än andra däggdjur med samma kroppsvikt. Detta gäller även människan. Men vi får en annan bild, om vi, i stället för basalmetabolismen, mäter den totala energiomsättningen i kilokalorier per minut. Den totala energiomsättningen är ett djurs energiomsättning under en längre tid, då det beter sig som det gör i naturen. I naturen vilar djuret inte hela tiden. Det förflyttar sig, äter och gör en mängd andra saker. Det visar sig att primater har en betydligt lägre total energiomsättning mätt per kilo kroppsvikt än de flesta andra däggdjur. Detta skulle kunna vara en förklaring till att primaterna, inklusive människan, lever längre än de flesta andra däggdjur med hänsyn tagen till kroppsvikten. En lägre total energiomsättning skulle ju kunna innebära att de är mindre utsatta för fria radikaler. Men detta har inte bevisats och det kan finnas andra förklaringar.
Sköldpaddor har väldigt låg energiomsättning. De behöver därför inte mycket mat och kan klara sig relativt länge utan mat. Sköldpaddor kan ju också bli mycket gamla. Kanske har deras höga ålder något med deras låga energiomsättning att göra. Men vi kan inte vara säker på att det finns ett orsakssammanhang och det finns många djur som lever längre eller kortare tid än vi skulle förvänta sig utifrån deras energiomsättning.
Skador av fria radikaler drabbar särskilt mitokondrierna, cellernas "kraftverk", som ombesörjer den aeroba cellandningen under förbrukning av syre. Skador på cellernas mitokondrier har antagits vara en viktig orsak till åldrande. Men även här är forskningsläget oklart.
Kan mTOR och sirtuiner ha betydelse för cellers åldrande?
Enligt en teori är åldrande kopplat till aktiviteten i den så kallade TOR-signaleringen inuti cellerna, hos däggdjur mTOR. Proteinet TOR bildar navet i ett mycket komplicerat nätverk, i vilket ett flertal molekyler skickar signaler genom att påverka varandra. Nätverket påverkas både av stimuli utifrån till cellerna och stimuli inuti cellerna. Det reglerar en rad processer i cellen, bland annat ämnesomsättning, tillväxt, celldelning och cellens överlevnad. Särskilt intressant är att ämnet rapamycin, som hämmar TOR-signalering, har visat sig öka livslängden hos flera djurarter, bland annat möss. Men mycket är oklart inom detta område och mer forskning behövs.
Sirtuiner är en grupp av enzymer som tros ha betydelse för åldrande. Resveratrol, ett ämne som aktiverar ett sirtuin, anses motverka åldrande. Men även här är forskningläget oklart.

Kan glukos bidra till åldrande?
En annan faktor som kan bidra till åldrande är att glukos, som vi får i oss i födan eller bildar själva, reagerar med och binds till långlivade stora molekyler, till exempel proteiner. Detta kan negativt påverka molekylernas funktion och därmed funktionen hos celler och vävnader. Sådan glykering av proteiner anses bland annat vara en av de faktorer som ger upphov till grumling av ögats lins vid grå starr (katarakt).
Det kan vara värt att nämna att läkemedlet metformin har visat sig förlänga livet hos vissa försöksdjur. Metformin användes vid diabetes typ 2 ("åldersdiabetes"). Det sänker blodets glukoshalt via flera mekanismer, bland annat genom att öka vävnadernas insulinkänslighet så att deras glukosupptag ökar, genom att minska leverns glukosproduktion och genom att fördröja absorptionen av glukos i tarmen. Forskarna vet emellertid inte hur metformin förhindrar åldrande.
Kan hormonet IGF-1 bidra till åldrande?
IGF-1 ("insulinlike growth factor 1") är ett hormon som fungerar som tillväxtfaktor bland annat genom att stimulera cellers tillväxt och utveckling. Hos däggdjur bildas IGF-1 bland annat i levern, där dess insöndring stimuleras av tillväxthormon (GH). Hos rundmasken Caenorhabditis elegans, bananflugor och möss har forskarna ökat djurens livslängd genom att minska halten av IGF-1 i kroppen. Men brist på iGF-1 kan också orsaka sjukdomar hos däggdjur. Dess betydelse för åldrandet hos människan är därför oklar.
Kan skadliga inflammationer orsaka åldrande?
Det har också föreslagits att kroppens inflammatoriska system uppregleras vid åldrande, så att vi ständigt bär på milda, men i längden skadliga, inflammationer. Inflammationer är i och för sig ett av kroppens försvarssystem mot infektioner och skador. Men ofunktionella inflammationer anses bidra till en rad sjukdomar, som åtföljer åldrandet, till exempel hjärt-kärlsjukdom, diabetes och Alzheimers sjukdom.
Kan förkortade kromosomändar döda celler?
Ytterligare en faktor som kopplats till åldrande är längden av de så kallade telomererna. Telomererna är DNA-sekvenser som skyddar ändarna på kromosomerna. När kromosomantalet fördubblas i samband med att kroppsceller delar sig (så kallad mitos) så startar DNA-syntesen vid varje kromosom en bit från kromosomänden och en del DNA går förlorat. Telomererna säkerställer att inga informationsbärande gener går förlorade vid DNA-syntesen. Men telomererna blir kortare vid varje delning och begränsar därmed antalet delningar som cellen kan genomgå. När telomererna är förbrukade leder kromosomabnormiteter till att cellen dör. Telomerernas roll är komplicerad, men det finns data som tyder på att människor med längre telomerer tenderar att leva något längre.
Kan mutationer i kroppscellerna leda till åldrande?
En orsak till åldrande skulle kunna vara DNA-skador, som orsakar mutationer i kroppens celler. Det finns flera enzymsystem som reparerar DNA. Men de lagar inte alla skador. Mutationer i cellerna kan ge upphov till cancer och tros även påskynda åldrande. Fladdermöss lever mycket längre än vi skulle förvänta sig för så små djur. Detta skulle bland annat kunna bero på att de har en ovanligt bra förmåga att reparera sitt DNA.
Kan förlust av stamceller begränsa livlängden?
Ännu en orsak till åldrande kan vara att stamcellerna blir färre eller slutar att fungera normalt. Vuxna stamceller är celler som finns i relativt litet antal i kroppens vävnader. De kan dela sig och ger då upphov dels till nya stamceller, dels till celler som kan differentieras och förändras till normala vävnadsceller. Stamceller är särskilt aktiva i vävnader där cellerna ständig bildas eller förnyas, till exempel överhuden, magtarmkanalens innersta cellager (epitelet) och den röda benmärgen. Blodet är en flytande vävnad som innehåller celler. Så kallade hematopoetiska stamceller i den röda benmärgen ger i flera steg ständigt upphov till nya röda blodkroppar, vita blodkroppar och blodplättar, allteftersom dessa cellulära beståndsdelar i blodet dör. Hos en 115-årig kvinna fann forskare att två tredjedelar av hennes vita blodkroppar härstammade från bara två stamceller, vilket tyder på att nästan alla hennes hematopoetiska stamceller dött. Skelettmusklernas stamceller, satellitcellerna, kan bland annat ge upphov till nya muskelfibrer i skadade muskler och vid styrketräning. Det finns data som tyder på att satellitcellerna fungerar allt sämre ju äldre vi blir.
Har naturligt urval ökat människans livsängd?
Den välutvecklade intelligensen och förmågan att planera för framtiden kan ha skyddat de tidiga människorna från rovdjur och födobrist. Hos människor, precis som hos sköldpaddor, skulle det därför kunnat ha varit en fördel att genom evolutionär anpassning öka livslängden. En möjlig förklaring till människans långa liv är således att hon löper mindre risk att dödas av rovdjur, tack vare sin kroppstorlek, sitt välutvecklade sociala liv och sin avancerade kultur. Då kan det vara en evolutionär fördel att fortplanta sig under flera år och att producera en liten avkomma som vårdas under lång tid.
En extra fördel med ett långt liv för oss människor är att det ges tid att överföra det kulturella arvet från generation till generation. Därmed hinner varje generation med att lära nästa generation en mängd saker som ökar överlevnadsförmågan. Det behövs, med andra ord, ordentlig utbildning för att lära sig tillverka stenyxor, tända eld, samla ätliga rötter och skjuta med pilbåge.
Summering
Det finns med största sannolikhet inte en orsak till åldrandet. I stället leder troligen ett komplicerat samspel mellan flera av de ovan nämnda och andra mekanismer till att kroppens funktioner successivt försämras.
Fördjupning
- Världens äldsta djur: om olika organismers livslängd. Finns det djur som inte åldras, undflyr döden och kan leva ett evigt liv? Du hittar artikeln under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
- Om möss och elefanter: kroppsstorlekens betydelse för djurs ämnesomsättning. Artikeln finns under rubriken Varför är vissa djur stora medan andra är små?.
- Hur nya ben och svansar ersätter gamla. Regeneration: återväxt av amputerade kroppsdelar. Du kan läsa artikeln under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?.
Referenser
- E. Barbi et al.: The plateau of human mortality: demography of longevity pioneers (Science 360:1459-1461, 2018).
- G. Bjelakovic et al.: JAMA clinical evidence synopsis. Antioxidant supplements to prevent mortality (JAMA 310:1178-1179, 2013).
- H. M. Blau et al.: The central role of muscle stem cells in regenerative failure with aging (Nature Medicine 21:854-861, 2015).
- S. Chang: Modeling aging and cancer in the telomerase knockout mouse (Mutation Research 576:39-53, 2005).
- X. Dong, B. Milholland, och J. Vijg: Evidence for a limit to human lifespan (Nature, publicerad online 5 october 2016).
- R. Doonan et al.: Against the oxidative damage theory of aging. Superoxide dismutases protect against oxidative stress but have little or no effect on life span in Caenorhabditis elegans (Genes & Development 22:3236-3241, 2008).
- N. Fleming: The tricks that help some animals live for centuries (BBC Earth, uppdaterad 31 mars 2017).
- D. Gabuzda och B: A. Yankner: Inflammation links ageing to the brain (Nature 497:197-198, 2013).
- L. Geddes: Human age limit claim sparks debate (Nature News, 5 october 2016)
- J. Gil and D. J. Withers: Out with the old (Nature 530:164-165, 2016).
- D. E. Harrison et al.: Rapamycin fed late in life extends lifespan in genetically heterogeneous mice (Nature 460:392-396, 2009).
- Ö. Hemström: Medellivslängden ökar stadigt (Statistiska centralbyrån, publicerat den 3 oktober 2012).
- H. Holstege et al.: Somatic mutations found in the healthy blood compartment of a 115-yr-old woman demonstrate oligoclonal hematopoiesis (Genome Research 24:733-742, 2014).
- D. Holmes och K. Martin: A bird's-eye view of aging. What's in it for ornithologists (The Auk 126:1-23, 2009).
- J. P. Kehrer och L.-O. Klotz: Free radicals and related reactive species as mediators of tissue injury and disease, implications for Health (Critical Reviews in Toxicology 45:765-798, 2015).
- S. J. Olshansky: Measuring our narrow strip of life (Nature, publicerad online 5 october 2016).
- V. I. Pérez et al.: The overexpression of major antioxidant enzymes does not extend the lifespan of mice (Aging Cell 8:73-75, 2009).
- M. Prokocimer, R. Barkan, och Y. Gruenbaum: Hutchinson–Gilford progeria syndrome through the lens of transcription (Aging Cell 12:533-543, 2013).
- M. Ristowa et al.: Antioxidants prevent health-promoting effects of physical exercise in humans (Proceedings of the National Academy of Science 106:8665-8670,2009).
- M. Scudellari: The science myths that will not die (Nature 528:322-325, 2015).
- Socialstyrelsen: Folkhälsa (uppdaterat den 22 februari 2016).
- B. J. Willcox et al.: Caloric restriction, the traditional Okinawan diet, and healthy aging (Annals of the New York Academy of Science 1114: 434-455, 2007).

Äkta löss (Anoplura) är alla blodsugare på däggdjur. De är insekter som förlorat vingarna. Det finns två underarter av människolusen Pediculus humanus, klädlusen och huvudlusen, ibland betraktas de som två åtskilda arter. Klädlusen är numera ovanlig i Sverige, medan huvudlusen fortfarande förekommer. Även flatlusen (Pthirus pubis), som är en särskild art, förekommer i Sverige. Den lever främst kring könsorganen, men också på andra hudområden där det finns grova hårstrån.
Lusen på bilden ovan har på huvudet två korta antenner. På mellankroppen har den sex gångben med kraftiga klor som används när den klamrar sig fast vid håret eller kläderna. Bakkroppen består av många segment.
Studier av DNA tyder på att huvudlus och klädlus skildes åt för mellan 170 000 och 83 000 år sedan. Resultaten tolkades som att den moderna människan (Homo sapiens) vid denna tidpunkt började bära kläder, förmodligen i form av djurhudar. Detta skedde innan hon utvandrade från Afrika till resten av världen. Intressant nog inleddes en istid för mellan 190 000 och 130 000 år sedan. Det kyligare klimatet drabbade även Afrika och kan ha varit orsaken till att människorna där behövde kläder. Kläderna kan ha gynnat den moderna människans spridning ut ur Afrika till kyligare områden. Vi vet inte om neandertalmänniskan (Homo neanderthalensis) använde kläder, men klart är att hon genomlevde en tidigare istid i Europa. Det är svårt att tro att hon hade klarat sig utan kläder. I så fall var den moderna människan troligen inte den första människoart som ekiperade sig.
Forskare har också jämfört DNA hos gorillans och människans flatlus. En tolkning av resultaten är att kroppsbehåringen förlorades hos en förfader till den moderna människan redan för 3-4 millioner år sedan, således långt innan kläder började användas.
Referenser
- R. Dunn: Of lice and men: A very intimate history (New Scientist, 6 November 2012).
- D. L. Reed, J. E. Light, J. M. Allen, och J. J. Kirchman: Pair of lice lost or parasites regained: the evolutionary history of anthropoid primate lice (BMC Biology 5:7, 2007).
- M. A. Toups, A. Kitchen, J. E. Light, och D. L. Reed: Origin of clothing lice indicates early clothing use by anatomically modern humans in Africa (Molecular Biology and Evolution 28:29–32, 2011).

Varför har giraffen så lång hals?
Giraffen har, som bekant, den längsta halsen bland nu levande ryggradsdjur. Det finns många svar på den frågan. För att den ska kunna beta högt upp i trädkronorna? Det är inget bra svar. Giraffen har inte konstruerats med syftet att göra detta. Ett bra svar är att beskriva de mekanismer som gör att giraffens hals tillväxer, så att den blir den längsta halsen bland nu levande djur. Ett helt annat och lika bra svar är att den långa halsen har uppkommit genom flera generationers evolution.
Men vad innebär evolution? Jean-Baptiste de Lamarck (1744–1829) hävdade, mycket förenklat, att giraffer som betade i trädkronor fick längre halsar och att denna förvärvade egenskap gick i arv till nästa generation. Efter flera generationer hade girafferna fått mycket långa halsar. Vi vet nu att evolutionen inte fungerar på det viset. Lamarck var emellertid en stor vetenskapsman. Han vann inte gehör i sin samtid, men banade väg för Charles Darwins evolutionslära, en av de bästa och mest fruktbärande vetenskapliga teorierna någonsin.
Enligt Darwins evolutionslära har giraffen fått sin långa hals genom slumpvisa genetiska mutationer och naturligt urval (naturlig selektion). Giraffindivider med en mutation, som förlängde halsen eller benen, fick tillgång till mer föda. Detta berodde på att dessa individer kunde beta blad högre upp på träden, dit inga andra djur nådde. Därigenom ökade deras livsduglighet och de fick en större avkomma än korthalsade individer. De gynnades med andra ord av det naturliga urvalet. Flera mutationer, som förlängde halsen och benen, ökade så småningom i frekvens, eftersom bärare av dessa mutationer fick fler giraffkalvar än andra giraffer. Så småningom eliminerades de genvarianter som gav kort hals och korta ben helt från giraffens arvsmassa.
Giraffen har förmodligen ytterligare en fördel med sin långa hals och sina långa ben. När vinder blåser runt smalare cylindriska utskott, som hals och ben, har den en större avkylande effekt, än när den blåser runt den tjockare kroppen. Mer värme avges då genom direkt värmetransport till luften och genom avdunstning av vatten från huden. Giraffen långa hals och ben skulle alltså kunna skydda den mot överhettning på den varma solbelysta savannen. Den viktigaste fördelen med giraffens hals och ben förblir dock ändå förmågan att beta högt upp i träden
Molekylärbiologer har identifierat 70 genvarianter, som finns hos giraffen, men inte hos andra däggdjur, inklusive giraffens närmaste släkting, okapin. Flera av dessa genvarianter har betydelse för utvecklingen av halsens och benens skelett. En del av dem är så kallade Hox-gener, gener som styr kroppens utformning under fosterutvecklingen. Andra genvarianter styr hjärt-kärlsystemets utveckling. Det är sannolikt dessa genvarianter, som gynnades av det naturliga urvalet, när giraffens hals och ben förlängdes. Mutationerna i gener, som styr hjärt-kärlsystemets funktion, behövdes för att motverka de cirkulatoriska problem som den långa halsen och de långa benen ger upphov till på grund av tyngdkraften.
Ordet "varför" uttrycker en fråga. Vi frågar efter orsaken till något. I dagligt tal handlar det ofta om avsikt, om att någon människa har utfört en handling med ett syfte eller ett mål. Det är därför lätt att missuppfatta ordet "varför" i vetenskapliga sammanhang. Nedan diskuteras tre betydelser som detta svårtolkade ord kan ha.
Teleologisk orsak
Detta innebär en framförliggande, målinriktad orsak och är inte förenligt med naturvetenskapen. Vilket mål har funnits med ett djur? Aristoteles ansåg, teleologiskt, att röken strävar uppåt därför att den hör hemma där, det vill säga att dess mål finns där uppe. Företrädare för så kallad "intelligent design" anser att en skapare har gjort en "ritning" för ett djur, alltså ställt upp ett mål, och sedan konstruerat det. Den katolske prästen Pierre Teilhard de Chardin (1881-1955) var en skicklig paleontolog som accepterade evolutionen, men inte darwinismen. Han ansåg, teleologiskt, att evolutionen strävade mot ett mål, Omega. Han ansåg att Omega var ett gudomligt tillstånd av maximal komplexitet och medvetande. Han ansåg att hela universum strävade emot Omega.
Vi förkastar teleologiska resonemang, eftersom det inte finns någon som helst evidens för dem i naturvetenskapliga sammanhang. Då återstår två naturvetenskapliga, bakomliggande orsaksdefinitioner.

Proximat orsak
Hur är ett djur uppbyggt och hur fungerar det? Vilka bakomliggande mekanismer, i form av orsakskedjor och orsaksnätverk, förklarar att djuret ser ut och fungerar som det gör? Vilka är de komplicerade bakomliggande orsaker som lett till att dess kropp uppkommit ur ett befruktat ägg under embryots utveckling? Vilka bakomliggande mekanismer gör att djurets hjärta slår, att det kan röra sig och att det kan tillgodogöra sig sin föda?
Låt oss ta komodovaranen, världens största ödla, som ett exempel. Detta djur har ett bra luktsinne och kan röra sig snabbt över korta sträckor. De proximata orsakerna till det goda luktsinnet skulle bland annat kunna vara att varanen har mycket känsliga luktreceptorer och att den är utrustad med ett stort antal olika luktreceptorer för olika dofter. De proximata orsakerna till varanens rörelseförmåga har att göra med musklernas egenskaper och rörelseapparatens uppbyggnad.
Vilken bakomliggande orsakskedja gör att röken stiger uppåt? Så här är det. Elden värmer den luft som innehåller röken. Luftmolekylerna får då ett högre energiinnehåll. Därför rör de sig mer och kolliderar oftare med varandra. Det blir då glesare mellan luftmolekylerna. Eftersom avståndet mellan luftmolekylerna ökat, får den varma luften en lägre densitet ("täthet") än den omgivande kalla luften. Därför stiger röken uppåt buren av den varma luften.

Ultimat eller evolutionär orsak
Genom vilka bakomliggande mekanismer har ett djur uppkommit under evolutionen? Här har både slumpen och det naturliga urvalet medverkat, samt dessutom sexuellt urval. Slumpvisa mutationer har uppkommit i djurets arvsmassa, i enskilda gener eller genom förändringar på kromosomal nivå. Dessutom har befintliga gener stuvats om vid den könliga fortplantningen. Båda mekanismerna har lett till att individer med nya genkombinationer uppstått. Vissa individer har därigenom blivit bättre genetiskt anpassade, adapterade. till sin miljö, andra har blivit sämre adapterade. De bättre adapterade individerna kan till exempel ha blivit snabbare eller fått ett bättre luktsinne, vilket underlättat bytesfångsten. De mera väladapterade individerna är friskare och starkare. Därför har de fått en större avkomma. Då har deras genvarianter och genkombinationer ökat i frekvens i populationen, medan de mindre gynnsamma genvarianterna och genkombinationerna har försvunnit.
Observera att det naturliga urvalet inte är målinriktat. Det verkar här och nu, alltså vid den tidpunkt och i den miljö djuren befunnit sig vid det tillfälle då deras arvsmassa har förändrats. Richard Dawkins har formulerat detta i boktiteln "The blind watchmaker". Observera också att miljön innefattar både den icke levande omgivningen och andra levande organismer i ekosystemet.
Slumpen kan påverka evolutionen på ett annat sätt än genom slumpvisa mutationer. Små djurpopulationer kan ha isolerats från sina artfränder (genetisk drift). Alternativt kan små populationer ha utvandrat till ett nytt område (grundareffekt). I båda fallen kan genom slumpens inverkan vissa gener eller genkombinationer ha saknats i de nya populationerna. De nya populationerna kan med tiden ha utvecklats till nya arter. Detta kan kanske ha hänt när komodovaranen isolerats på öar som uppstod, när det som en gång var en sammanhängande landmassa omvandlades till dagens indonesiska övärld.
Ytterligare en synpunkt. Till skillnad från så kallad "intelligent design", leder naturligt urval inte till perfekta organismer, bara till organismer som överlever. En anledning kan vara att gynnsamma gener försvunnit i små populationer, som beskrivits ovan. En annan anledning kan vara att adaptioner till en miljö som djuret en gång levt i finns kvar, men inte längre är funktionella i den nya miljön. Ytterligare en anledning kan vara att det finns restriktioner i djurets fosterutveckling som gör att den vuxna individen inte kan bli perfekt adapterad.
Om hur förvärvade egenskaper i vissa fall kan ärvas
Lamarck har numera fått en viss revansch. Vi vet nu att förvärvade egenskaper i vissa fall kan ärvas. Det handlar då inte om förändringar i den genetiska koden i arvsmassans DNA. I stället rör det sig om att gener stängs av eller aktiveras genom att vissa molekyler binds till eller avlägsnas från arvsmassan, bland annat genom så kallad metylering eller avmetylering av DNA eller av kromosomala proteiner. I vissa fall kan dessa så kallade epigenetiska förändringar överföras till nästa generation. När dräktiga möss utsattes för vissa miljögifter, fann forskarna ökad sjuklighet i de följande fyra generationerna. Dessa sjukdomar troddes vara orsakade av nedärvda epigenetiska förändringar. Doserna av miljögifterna var dock betydligt högre än de som människor utsätts för i förorenade miljöer. Det finns studier av människor som tolkats så att svältperioder under barndomen skulle kunna leda till ökad sjuklighet hos barnbarnen. Möjligen skulle detta kunna bero på epigenetisk nedärvning, men här behövs mer data. Dessa resultat innebär dock inte att förvärvade egenskaper normalt ärvs. Sådan nedärvning måste betraktas som ovanlig. Det är fortfarande Darwins evolutionsteori som gäller, inte Lamarcks.
Fördjupning
Varifrån kommer vi? Om lansettfiskar, ryggradsdjur och människans ursprung. Du hittar artikeln under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
Referenser
- M. Agaba et al.: Giraffe genome sequence reveals clues to its unique morphology and physiology (Nature Communications 7:11519, 2016).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012) och en mängd andra biologiska läroböcker på universitetsnivå.
- M. K. Skinner: A new kind of inheritance (Scientific American 311:44-51, 2014).

Flugor är kända för att fortplanta sig snabbt. Hur många flugor skulle en flughona kunna ge upphov till under loppet av ett år? Naturligtvis väldigt många. Men räknar vi på det blir det ofattbart många fler flugor, än vi någonsin skulle kunna föreställa oss Vi förutsätter att alla flugor överlever så länge att de hinner med att ge upphov till största möjliga avkomma. I detta teoretiska flugparadis lever flugorna i den bästa möjliga miljön och har alltid gott om föda. Där finns inga insektsätande djur och flugorna drabbas inte av parasiter eller sjukdomar.
Låt oss då räkna på det, så får vi se. Vi väljer den vanliga husflugan (Musca domestica) som exempel. Uppgifterna om husflugan skiljer sig i de källor som jag utnyttjat. Dessutom det varierar många av mätvärdena med en rad olika faktorer, till exempel temperatur och olika flugpopulationers genetik. Slutresultatet blir således osäkert, men ger ändå en skrämmande klar uppfattning om antalet flugor.
En vuxen husfluga uppges i olika källor vara mellan 6 och 9 millimeter lång. Vi räknar med 7 millimeter. Flugorna måste ha ståplats. Jag har med hjälp av bilder skattat avståndet mellan de yttersta fötterna och kommit fram till att en fluga som sitter på ett vågrät plan upptar en yta på cirka 35 kvadratmillimeter. Om flugan är 3 millimeter hög upptar den en volym på 105 kubikmillimeter.
En flughona uppges kunna lägga maximalt cirka 500–600 ägg under sin livstid. Vi räknar med 500 ägg per hona.
Det maximala antalet fluggenerationer under ett år uppges vara mer än 20. Detta gäller i subtropiska och tropiska områden, där flugorna kan föröka sig hela året. Vi räknar med 20 generationer per år.
Vi förutsätter sedan att alla flugor överlever från ägg till könsmogen fluga och att varje flughona lägger 500 befruktade ägg, som alla kläcks och ger upphov till nya flugor. Vi förutsätter också att alla flugorna dör omedelbart efter äggläggningen. Notera att cirka hälften av äggen ger upphov till hanar, cirka hälften till honor.
Om vi börjar med en befruktad flughona, som lägger 500 ägg från och med nyårsdagen och alla äggen utvecklas till könsmogna flugor får vi 250 nya honor i det nya årets första generation. I den andra generationen får vi 62 500 honor, det vill säga 250 x 250 eller 2502 honor. I den tredje generationen får vi 15 625 000 honor, det vill säga 15,6 miljoner, 250 x 250 x 250 eller 2503 honor. I den 20:e generationen vid årets slut får vi 25020 honor, vilket motsvarar 9 x 1047 eller
900 000 000 000 000 000 000 000 000 000 000 000 000 000 000 000 honor
Notera att varje generation, förutom honorna, innehåller ungefär lika många hanar. Totalt får vi således 18 x 1047 eller
1 800 000 000 000 000 000 000 000 000 000 000 000 000 000 000 000 flugor
Om y är antalet honor i varje generation och x antalet generationer har vi en så kallad exponentialekvation: y= 250x. Exponentiell tillväxt hos organismer ger som synes en ofantligt snabb ökning i individantal.
Om 18 x 1047 flugor, hälften honor och hälften hanar, står bredvid varandra så upptar de alla tillsammans en yta på cirka 6 x 1037 kvadratkilometer. Deras sammanlagda volym är cirka 2 x 1032 kubikkilometer. De får inte plats på jordens yta som bara är cirka 5,1 x 108 kvadratkilometer och inte heller på solens yta som är cirka 6,1 x 1012 kvadratkilometer. De får inte ens plats inom solens volym som är cirka 1,4 x 1018 kubikkilometer.
I verkligheten kläcks lyckligtvis inte alla ägg och det överväldigande flertalet larver, puppor och flugor dukar lyckligtvis under för insektsätande djur, parasiter och sjukdomar, innan de har hunnit fortplanta sig. Ytterst få flugor lever så länge att de dör av ålderdomssvaghet efter cirka 3-4 veckor som vuxna. Därmed hålls jordens flugpopulation inom rimliga gränser. Och det naturliga urvalet sållar bort de sämre anpassade och mindre fortplantningsdugliga flugorna.
Flugornas evolutionära strategi är att producera en mycket stor avkomma utan att ha någon yngelvård, så kallad r-selektion.
Referenser
- W. Iqbal et al.: Role of housefly (Musca domestica, Diptera; Muscidae) as a disease vector, a review (Journal of Entomology and Zoology Studies 2:159-163, 2014).
- Musca domestica (Encyclopedia of Life, hämtad 15 april 2016).
- Musca domestica Linnaeus (Entomology & Nematology, University of Florida, hämtad 15 april 2016).
- J. A. Rozendaal: Houseflies. Carriers of diarrhoeal diseases and skin and eye infections (pp. 302-323 in J. A. Rozendaal "Vector control: methods for use by individuals and communities", World Health Organization, 1997).
- Solar system exploration: Earth (NASA, hämtad 15 april 2016).
- Solar system exploration: Sun (NASA, hämtad 15 april 2016).
Märkliga och farliga djur
Elefanter dricker som bekant genom att suga upp vatten i snabeln och sedan spruta in det i munnen. Det ser mycket enkelt ut. Men i själva verket är det en märklig prestation, som forskare inte har kunnat förklara.
Elefanternas uppsugning av vatten i snabeln är ett mycket intressant problem. För något år sedan kontaktades jag av en lungläkare, som skrev en artikel om elefanternas lungor. Vi diskuterade problemet och letade i litteraturen, men fann inget bra svar. Forskarna har inte gjort lämpliga försök på elefanter. Därför finns det bara obestyrkta hypoteser.
Problemen med att ta upp vatten i snabeln
En möjlighet är att elefanten utvidgar lungorna, som vid en inandning. Då drar den ihop mellangärdesmuskeln så att den rör sig bakåt, varvid lungorna utvidgas, så att ett undertryck uppstår i dem relativt atmosfärstrycket. I så fall skulle lungorna fungera som en sugpump, som drar upp vatten i snabeln. Röstspringan (glottis) i struphuvudet måste då vara öppen och strupplocket (epiglottis) uppfällt, så att luft kan komma in i lungorna. Risken finns då att vatten kommer ner i lungorna. Vid sväljning stängs dessa strukturer, så att mat och dryck inte hamnar i lungorna. Betänk dock att snabeln hålls lodrät. Med en finstämd kontroll av snabelmuskulaturen skulle elefanten kunna manövrera så att vatttenytan inte når över snabelhålrummets övre kant och därmed inte hamnar i lungorna.
Intressant är att elefanter kan suga upp vatten med öppen mun, något som vi skulle tro vara omöjligt. Men de har, till skillnad från de flesta andra däggdjur, en slutmuskel som kan stänga förbindelsen mellan munhålan och svalget. När denna förbindelse är stängd, är det bara snabeln som via näshålan har förbindelse med lungorna.
Det finns dock ett annat och betydligt värre problem med att använda lungorna som sugpump. De lodräta snabeln är cirka 2 meter lång. Om lungorna ska suga upp vatten 2 meter, måste lufttrycket i dem var cirka 20 procent lägre än atmosfärstrycket, extremt lågt jämförbart med vid en vanlig inandning. Då blir trycket i lungornas blodkapillärer extremt mycket högre än i den angränsande lungluften. Det borde finnas en uppenbar risk att kapillärerna spricker så att lungorna fylls med blod. Några egenheter i lungornas anatomi skulle möjligen kunna motverka detta.
Notera dock att elefanten inte suger i sig vatten enkelriktat genom hela snabeln som vore den ett sugrör. Den suger upp vatten en bit upp i snabeln och sprutar sedan in det i munnen. Men även om den bara suger upp vatten en halv meter, måste trycket i lungorna vara cirka 5 procent lägre än atmosfärstrycket, ett undertryck på cirka 40 mm Hg. Det ovan diskuterade problemet med lungkapillärerna kvarstår även då.
Möjliga lösningar på elefantens problem
De tillplattade lungsäckarna, belägna mellan brösthålans väggar och lungorna, är hos elefanter inte fyllda med vätska, som hos andra däggdjur, utan med bindvävnad. Lungsäckarnas tunna väggar är förstärkta med bindvävnad. Bindväven förhindrar kanske att lungsäckarnas kapillärer brister. Inuti lungorna är blodkärlen omgivna av förstärkande bindväv. Men detta gäller inte de tunnaste blodkärlen, de gasutbytande kapillärerna, som är belägna tätt intill de gasfyllda lungblåsorna (alveolerna). Så problemet kvarstår. Med lungorna som sugpump borde rimligen lungkapillärerna brista. En möjlighet är dock att kapillärerna utsätts för tryckstress under så pass kort tid att de inte hinner brista.
Elefanter är försedda med en säckliknande utbuktning från svalgets golv. En hypotes är att de suger in vatten i snabeln med hjälp av muskler, som utvidgar denna svalgsäck. Då skulle vägen till lungorna kunna vara stängd. Men säckens volym verkar vara för liten och forskarna har inte hittat några muskler som skulle kunna utföra arbetet.
Ytterligare en hypotes är att de suger upp vatten i snabeln med hjälp av dess muskulatur, antingen genom att utvidga dess inre hålrum eller genom någon slags peristaltik. Något stöd för denna hypotes tycks inte finnas. Peristaltik är den mekanism genom vilken födan transporteras av vandrande muskelsammandragningar genom tarmen.
Frustrerad och dyster tittade jag på ett antal YouTube-videor med drickande elefanter. Men när djuren sög upp vatten, kunde jag inte iakttaga någon volymökning, varken i bålen, svalgregionen eller snabeln. Mysteriet förblir olöst.
Elefanter härstammar troligen från djur som delvis levde i vatten. De är faktiskt duktiga simmare. På grund av sin tunga benstomme ligger de då lågt i vattnet. Men de löser problemet genom att använda snabeln som snorkel. Men även detta leder till problem som ännu inte har utretts.
Fördjupning
Om möss och elefanter: kroppsstorlekens betydelse för djurs ämnesomsättning. Du hittar artikeln under rubriken Varför är vissa djur stora medan andra är små?
Referenser
- R.E. Brown et al.: The elephant’s respiratory system: adaptations to gravitational stress (Respiration Physiology 109:177-194, 1997).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012 ).
The peculiar lungs of the elephant (New Scentist, 6 december 1962). - J. B. West: Snorkel breathing in the elephant explains the unique anatomy of its pleura< (Respiration Physiology 126:1-8, 2001).

Alla känner till att flugor kan gå på taket. Mindre känt är att det finns ödlor som kan göra detsamma. Geckoödlornas tår kan fästa i nästan alla underlag, dock ej teflon. Och de fäster genom så kallad torr adhesion utan att vara klibbiga som tejp.
Många av våra tekniska framsteg realiserades genom evolution inom djurvärlden åtskilliga miljoner år innan de "uppfanns" av människan, bland annat hävstänger, brospann och värmeväxlare. Numera letar vi efter tekniska lösningar i djurvärlden och försöker imitera dem. Ett exempel på detta är geckoödlornas fantastiska fötter.
Geckoödlor kan gå på lodräta ytor. De kan till och med gå upp och ner, till exempel på ett tak, utan att ramla ner. De kan göra detta på många olika underlag: släta och skrovliga, torra och våta, vattenskyende och fettskyende. Redan den gamle greken Aristoteles funderade på hur de bär sig åt. I vår tid har en rad olika hypoteser framkastats. Fötterna är inte klibbiga och de är helt torra, så geckorna använder inte klister och utnyttjar inte heller vattnets kapillärkraft. Geckornas fötter fungerar inte som sugkoppar och inte som kardborrband. De verkar inte heller med hjälp av elektrostatiska krafter.
Vi vet nu att geckorna utnyttjar svaga så kallade van der Waals-krafter för att fästa i underlaget. Dessa krafter orsakas av att elektronmolnen mellan två atomer växelverkar så att de rör sig i takt med varandra. Detta innebär att molekyler på tårnas yta måste nå nästan direkt kontakt med molekyler i underlaget. Avståndet mellan små hudutskott på tårna och underlaget är då i samma storleksordning som mellan molekylerna i en kristall. Hur kan ödlorna åstadkomma detta?

Geckornas tår är på undersidan försedda med skivor, lameller. Vi trodde förr att lamellerna fungerade som sugkoppar. Men de gör de inte. Sugkoppar sitter fast på grund av att lufttrycket är lägre inne i sugkoppen än utanför. Eftersom geckotår fungerar också i vacuum, kan vi utesluta sugkoppshypotesen. För att komma ödlornas hemlighet på spåren måste vi se på tårnas yta i mycket hög förstoring. Det handlar om strukturer på huden som är mindre än en tusendels millimeter.

På undersidan av geckornas tår finns mycket små hårliknande utskott, setae, se bild ovan. På tokajgeckons tår finns det ungefär 14 400 setae per kvadratmillimeter. I de uppsplitsade ändarna på setae finns ett antal otroligt små spatulae, se bild ovan. Det är spatulae som fäster i underlaget. I och med att setae är böjliga, kommer ett enormt stort antal spatulae vidröra underlaget, även om detta är ojämnt.
Spatulae är inte klibbiga och fäster därför inte om de bara lätt vidrör underlaget. Om foten trycks mot underlaget, fäster de däremot med hjälp av de ovan nämnda van de Waals-krafterna, som binder molekyler i spatulae till molekyler i underlaget. En spatula binder med en mycket liten kraft, men eftersom det finns så enormt många bundna spatulae, så blir den totala kraften otroligt stor. Teoretiskt skulle en gecko kunna belastas med en vikt motsvarande två vuxna människor utan att den släpper taget från underlaget.
I praktiken är den totala kraften mindre, bland annat beroende på hur pass jämnt underlaget är. Men kraften är ändå tillräcklig för att en gecko ska kunna gå på taket och motstå en tropisk storm. Kraften är också tillräcklig för att geckon ska kunna få fotfäste i en gren, om den skulle råka falla, trots att den inte har gripförmåga. En fördel med att utnyttja van de Waals-krafter är att fäste kan åstadkommas på alla typer av underlag. Det krävs dock att avståndet mellan spatulae och underlaget blir så litet att van der Waals-krafterna kan verka. Men på grund av tåsulornas konstruktion åstadkommer geckorna tillräckligt nära kontakt på nästan alla underlag. Forskare har börjat undersöka den kemiska sammansättningen hos setae, men kan ännu inte förklara vad som ger dem så effektiv vidhäftningsförmåga.

Men hur kan en gecko övervinna tårnas oerhörda fästkraft, när den ska lyfta foten från underlaget? Svaret är att den, när den rör foten, först ändrar vinkeln mellan setae och underlaget utan att lösgöra tårna. Detta leder troligen till att bindningarna mellan spatulae och underlaget försvagas så att foten kan lyftas.
Trots att geckornas tåsulor kan binda tiill underlaget med så stor kraft. är de märkligt nog självrengörande. Tårnas undersidor är i och för sig starkt vattenskyende (hydrofoba) och fettvänliga (lipofila), men ingetting fastnar på dem, varken fettlöslig eller vattenlöslig smuts. Ödlorna behöver således inte rengöra fötterna. De lämnar inte heller någon smuts på underlaget, när de går. Tårnas setae har den egendomliga egenskapen att de bara fäster vid något om de utsätts för mekanisk deformering.
Utvecklare har naturligtvis arbetat med att framställa konstgjord tejp med samma egenskaper som geckornas tåsulor. Alla har vi råkat ut för att tejpen vi ska använda klibbar ihop när vi ska applicera den och att det blir märken på underlaget när vi tar bort tejp. Ingetdera av detta skulle hända med geckotejp. Geckotejp skulle dessutom vara så stark att den skulle kunna ersätta skruvar, spik och pluggar. Med geckotejp på handskar skulle fotbollsmålvakter bokstavligen kunna klistra bollar och bergsklättrare klättra på undersidan av bergsformationer. I båda fallen skulle de kunna lösgöra handskarna utan problem. Det har tagit relativt lång tid att utveckla geckoinspirerade material, men ett flertal geckoprodukter finns nu tillgängliga.
Fördjupning
Vilka djur hoppar högst? Hur kan små djur, som loppor och gräshoppor, hoppa så högt? Du hittar artikeln under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?.
Referenser
- L. Alibardi: Review. mapping proteins localized in adhesive setae of the tokay gecko and their possible influence on the mechanism of adhesion (Protoplasma 255:1785-1797, 2018).
- K. Autumn: How gecko toes stick (American Scientist 94:124-132, 2006).
- K. Autumn et al.: Evidence for van der Waals adhesion in gecko setae (Proceedings of the National Academy of Science USA 17:12252-12256, 2002).
- A. K. Geim et al.: Microfabricated adhesive mimicking gecko foot-hair (Nature Materials 2:461-3, 2003).
- A. Y. Stark et al.: The effect of temperature and humidity on adhesion of a gecko-inspired adhesive: implications for the natural system (Scientific Reports 6:30936, 2016).

Kamelfamiljen uppkom i Nordamerika för cirka 45 miljoner år sedan. Den spred sig för cirka fem miljoner år sedan till Asien vi en landbrygga över Berings sund och för cirka två miljoner år sedan till Sydamerika, när denna kontinent förbands med Nordamerika via den landbrygga som i dag är Mellanamerika.
Av de cirka 50 kameldjursarter vi damerika samt den tvåpuckliga eller baktriska kamelen (Camelus ferus) i Centralasien. Tamformen av tvåpucklig kamel benämns Camelus bactrianus. Den enpuckliga kamelen eller dromedaren (Camelus dromedarius) är utdöd som vild art. Som tam förekommer den främst i Mellanöstern, Nordafrika och västra Centralasien. Förvildade djur förekommer i Australien. Dagens kameldjur är specialiserade till att leva i torra miljöer (kamelerna) och på hög höjd (lamadjuren). Möjligen har de konkurrerats ut av idissslarna (oxdjur, får och getter, antiloper, hjortdjur med flera) i andra miljöer.
Kamelernas pucklar och uttorkningsproblem
Kamelernas pucklar innehålleri inte lagrat vatten att användas under ökenvandringar. De innehåller energiförråd i form av fett.

Enpuckliga och tvåpuckliga kameler
Den vilda tvåpuckliga kamelen är ovanlig och starkt hotad. Den enpuckliga kamelen är, som sagt, utdöd som vild art. Båda arterna är vanliga som husdjur. Den enpuckliga och den tvåpuckliga kamelen kan para sig och få fertil avkomma. Hybriden har en enda puckel, men denna är längre än dromedarens från huvud till svans räknat. Jag har sett namnet "tülu" användas för hybriden. Hybriden är större är föräldrarna och tycks därmed uppvisa så kallad heterosis ("hybrid vigour"). Med heterosis avses förbättrad vitalitet vid korsning av två linjer, särskilt om de är inavlade. Fertiliteten minskar emellertid i följande generationer, om vi fortsätter att korsa hybrider med en av moderarterna. Detta visar att det handlar om två olika kamelarter. De båda tama kamelformerna har därför också olika ursprung.
Dromedaren tämjdes troligen cirka 3 000–2 500 år före vår tideräkning i södra delen av den Arabiska halvön, den tvåpuckliga kamelen cirka 4 000–3 000 år före vår tideräkning i Kina eller Mongoliet. Båda arterna används som riddjur och lastdjur. De producerar en ull av hög kvalitet. De används också som mjölkdjur och köttdjur. De är av särskild betydelse i torra områden, där nötkreatur har svårt att klara sig.

De fyra lamadjursarterna
De två vilda lamadjursarterna, guanacon och vikunjan, är båda hotade och riskerar att dö ut. Två former av lamadjur finns bara som tamdjur och räknas ofta som särskilda arter. De domesticerades troligen cirka 4 000–3 500 år före vår tideräkning i Sydamerika. Laman (Lama glama) används som lastdjur och tros härstamma från guanacon. Alpackan (Vicugna pacos, Lama pacos) producerar en mycket fin ull och tros härstamma från vikunjan. Det har emellertid förekommit hybridiseringar, som försvårar tolkningen av tamformernas härstamning. Båda arterna används som köttdjur.

Märkliga blodkroppar och antikroppar
Kameldjuren utmärker sig genom att ha ovala röda blodkroppar, inte runda tillplattade som nästan alla andra däggdjur. Dromedarens röda blodkroppar kan, utan att de tar skada, krympa extremt mycket vid uttorkning och svälla extremt mycket vid vattenöverskott. Troliga orsaker till detta är att cellmembranet och zonen under detta är förstärkta och att hemoglobinet har speciella vattenbindande egenskaper. Möjligen underlättar de röda blodkropparnas ovala form deras passage genom blodkapillärerna. Dromedaren kan dessutom förlora mycket kroppsvatten, utan att blodvolymen minskar, något som skulle kunna leda till cirkulatorisk chock.
Kameldjuren utmärker sig också genom att i blodet ha en typ av antikroppar med bara två i stället för fyra aminosyrekedjor. Sådana antikroppar har kommit till användning inom medicinen.

Vilka tamdjur domesticerades i Amerika?
Den inhemska amerikanska befolkningen hade bara ett fåtal tamdjur, innan européerna anlände. De medförde hunden när de först anlände till Amerika. Möjligen importerade de höns från Oceanien. De tämjde själva bara ett fåtal inhemska djur: kalkonen, marsvinet, laman, alpackan, myskankan och gaddlösa bin. Detta beror troligen på att det finns en brist på lämpliga vilda djur att tämja i Amerika. Koschenillsköldlusen användes dessutom för att framställa textilfärg, men den kan knappast kallas ett tamdjur. Benjamin Franklin hävdade att kalkonen borde bli USAs nationaldjur, i stället för den vithövdade havsörnen, som inte drar sig för att äta as. Det är synd att det inte blev så.
Referenser
- P. Bogner: Osmotic and diffusive properties of intracellular water in camel erythrocytes. Effect of hemoglobin crowdedness (Cell Biology International 29:731e736, 2005).
- P. Bogner et al.: Augmented water binding and low cellular water content in erythrocytes of camel and camelids (Biophysical Journal 75:3085-3091, 1998).
- K. E. Conrath et al.: Emergence and evolution of functional heavy-chain antibodies in Camelidae (Developmental and Comparative Immunology 27:87-103, 2003).
- K. K. Hirst: Llamas and alpacas. The domestication history of camelids in South America (ThoughtCo, uppdaterad 3 april 2018).
- K. K. Hirst: The origin histories of dromedary and bactrian camels (ThoughtCo, uppdaterad 8 mars 2017).
- A. M. Lister: Remedies for windy camels (Nature 390:658-659, 1997).
- J. Lärn-Nilsson och R. Hall: Kameldjur (Nationalencyklopedin, hämtad den 28 maj 2018).
- G. M. O. Maloiy , B. M. Rugangazi, och M. F. Rowe: Energy expenditure during level locomotion in large desert ungulates: the one-humped camel and the domestic donkey (Journal of Zoology 277:248-255, 2009).
- R. Yagil, U. A. Sod-Moriah, och N. Meyerstein: Dehydration and camel blood. II. Shape, size, and concentration of red blood cells (American Journal of Physiology 226:301-304, 1974).
- M. K. Yousef, M. E. D. Webster och O. M. Yousef: Energy costs of walking in camels, Camelus dromedarius (Physiological Zoology 62:1080-1088, 1989).

Gökhonor lägger som bekant ägg i andra fåglars bo och låter dem föda upp en gökunge, så kallad boparasitism eller häckningsparasitism. Olika gökhonor är specialiserade på olika värdarter och lägger ägg som liknar värdartens, ett per bo. Hur kan gökar lägga ägg som ser olika ut? Varför acceperar värdföräldrarna gökägget och gökungen, som vore det deras egen avkomma?
Gökägget härmar värdartens ägg
Först om äggets utformning hos gökar med olika värdarter. Hos göken (Cuculus canorus) finns det olika grupper av honor, så kallade gentes (sing. gens), som som lägger sina ägg i bon hos olika värdarter. Varje gens producerar ägg, som i färg, mönster och form liknar värdartens, en form av så kallad mimikry. De flesta menar att skillnaderna mellan äggen hos olika gentes väsentligen orsakas av genetiska skillnader. Vidare menar forskarna att dessa skillnader bara ärvs från hona till hona och inte finns hos hanarna. Men underlaget för denna hypotes har hittills varit ganska svagt.

Hur väljer gökhonan bon att lägga ägg i?
Så till frågan om hur gökhonorna väljer ut ett bo för äggläggning. Det korta svaret är att vi inte kan ge något klart svar, mer forskning behövs. Det finns emellertid flera hypoteser och visst experimentellt stöd för flera av dem. Här följer några hypoteser. Valet skulle kunna bero på:
- Att ungen präglas på sina värdföräldrar, en klassisk form av inlärning hos fåglar. Detta ligger nära till hands, men det finns faktiskt data som talar emot denna hypotes. Ett problem är att denna prägling naturligtvis bara måste påverka äggläggning, inte parningsbeteende.
- Att de flyttande gökarna på våren återvänder till det geografiska område där de har kläckts. Detta skulle dock försvåra spridning av arten till andra områden där värdarten finns. Mekanismen lämnar dessutom stort utrymme för misstag, att ägg läggs i fel värdartsbo.
- Att gökhonan väljer ett visst habitat, till exempel lövskog eller buskage. Även då finns det en stor risk för misstag. Eventuellt skulle honan leta efter mer detaljerade kännetecken inom habitatet ("mikrohabitat").
- Att gökhonan upptäcker aktiva föräldrafåglar vid boet och kanske identifierar värdarten.
- Att gökhonan identifierar boets utseende hos värdarten.
- Att gökhonan känner igen värdartens ägg. Märkligt nog finns det data som talar emot denna näraliggande hypotes.
Valet skulle kunna ske genom en kombination av flera av de ovanstående mekanismerna så att honan successivt snävar in valmöjligheterna och slutligen hamnar rätt. Det är oklart i vad mån arv och inlärning ligger bakom bovalet.

En evolutionär kapplöpning mellan göken och värdarten
Det förkommer en evolutionär kapplöpning mellan göken och dess värdarter. För göken gäller det att producera ägg som är så lika värdartens som möjligt. För världarten gäller det att avlägsna gökägget. I bästa fall ser värden att gökägget ser annorlunda ut eller, mera sällan, att det tillkommit ett extra ägg. Då kastar det ut gökägget från boet. Orsaken till att värdföräldrarna accepterar gökungen är inte helt klarlagd. Möjligen kan förstagångsföräldar lättare acceptera gökungar. Å andra sidan finns det värdarter som kan kasta ut gökungar ur boet. Detta gäller dock andra gökarter än den som finns i Sverige.
Hur ärvs gökäggets färg och form?
Åter till en intressant fråga, nämligen äggens utformning hos olika gentes (se ovan). Om äggets färg och form ärvs både via hanar och via honor, skulle det kunna leda till artbildning. Om en hane då parar sig med en hona, som tillhör en annan gens, skulle honan producera intermediära ägg, som inte liknar någon av de två värdarternas. Sådana ägg skulle löpa större risk att elimineras av värdföräldrarna. Det naturliga urvalet skulle således starkt gynna parning mellan hanar och honor tillhörande samma gens. Detta skulle så småningom leda till att arten splittrades i flera arter.
Om nedärvningen hos gökens olika gentes, som de flesta menar, sker bara maternellt, alltså från hona till hona, behöver nya arter inte bildas. Indelningen i gentes gäller då bara honorna. Då spelar honans gens ingen roll vid parningen. En hane kan para sig med honor av flera olika gens utan att det påverkar avkommans överlevnad. Den allra största delen av arvsmassan ärvs via båda könen, inte bara maternellt, och kommer därför inte att påverkas av det naturliga urvalet avseende äggets utformning. Detta är en förklaring till att vi använder termen gentes, inte underarter.

Troligen nedärvning bara via honan
Hur kan då en egenskap ärvas strikt maternellt, bara från hona till hona? Hos däggdjur sker detta endast via mitokondrierna, de cellorganeller som sköter om cellandningen. Alla mitokondrier i ett embryo kommer nämligen från äggcellen, inga från spermien. Mitokondrierna innehåller en kromosom i form av en cirkulär DNA-molekyl. Hos däggdjur förekommer även strikt paternell nedärvning, bara från hane till hane. Hanarna har i cellkärnan en Y-kromosom och en X-kromosom, honorna två X-kromosomer. Gener på Y-kromosomen kommer därför nedärvas paternellt. Även hos fåglar förekommer maternell nedärvning via mitokondrier. Men hos dem har nästan alltid hanarna i cellkärnan två Z-kromosomer, honorna en W-kromosom och en Z-kromosom. Även gener på W-komosomen kommer således hos fåglarna att nedärvas strikt maternellt, bara från hona till hona.
År 2016 publicerades det första molekylärbiologiska beviset för att äggfärgen hos göken ärvs strikt maternellt. Forskare identiferade en cirka 2,6 miljoner år gammal gens, i vilken alla honor lade omönstrade blåa ägg. Denna linje skilde sig markant från andra gökar både avseende sitt mitokondriella DNA och sitt W-kromosomala DNA, men inte avseende övrigt DNA. Det troligaste är att gener i W-kromosomen bestämmer äggfärgen. Det kan inte uteslutas att nedärvning via hanar sker inom andra gentes, men detta måste nu betraktas som mindre sannolikt.
Referenser
- A. Antonov et al.: Are cuckoos maximizing egg mimicry by selecting host individuals with better matching egg phenotypes? (PLoS ONE 7:e31704, 2012).
- M. de L. Brooke och N. B. Davies: A failure to demonstrate host imprinting in the cuckoo (Cuculus canorus) and alternative hypotheses for the maintenance of egg mimicry (Ethology 89:154-166, 1991).
- M. I. Cherry, A. T. D. Bennett och C. Moskát: Do cuckoos choose nests of great reed warblers on the basis of host egg appearance? (Journal of Evolutionary Biology 20:1218-1222, 2007).
- N. B. Davies: Cuckoo adaptations: trickery and tuning (Journal of Zoology 284:1-14, 2011).
- N. Davies: The reed warbler and the cuckoo: an escalating game of trickery and defence (University of Cambridge, uppdaterad 22 februari 2016).
- F. Fossøy et al.: Ancient origin and maternal inheritance of blue cuckoo eggs (Nature Communications 7:10272, 2016).
- F. Fossøy et al.: Genetic differentiation among sympatric cuckoo host races: males matter (Proceedings of the Royal Society B 278:1639-1645, 2011).
- O. Krüger och M. Kolss: Modelling the evolution of common cuckoo host-races: speciation or genetic swamping? (Journal of Evolutionary Biology 26:2447-2457, 2013).
- M. Stevens: Bird brood parasitism (Current Biology 23:R909-R913, 2013).
- Y. Teuschl, B. Taborsky, och M. Taborsky: How do cuckoos find their hosts? The role of habitat imprinting (Animal Behaviour 56:1425-1433, 1998).
- C. Yang et al.: How cuckoos find and choose host nests for parasitism (Behavioral Ecology 28:859-865, 2017).

Pesten är en av de dödligaste sjukdomar som drabbat mänskligheten. Den mest kända epidemin är digerdöden på 1300-talet. Det sägs att pesten sprids med loppor från råttor, men det är inte hela sanningen. I gamla tider trodde man bland annat att pesten orsakades av "skämd luft". Först år 1894 identifierades pestbakterien (Yersinia pestis) av läkaren Alexandre Yersin.
Bilden ovan visar en fågelloppa. Det är alltså inte en människoloppa, men alla lopparter är mycket lika varandra. Loppor är insekter som förlorat vingarna. Alla vuxna loppor suger blod från däggdjur eller fåglar. Larverna lever av allehanda organiskt avfall, ofta i värddjurets bon, till exempel smuts i springor mellan golvbrädor i människans bostäder. Människoloppan, Pulex irritans, är numera sällsynt i Sverige. Dess minne har dock odödliggjorts i en dikt av skalden Nils Ferlin: "Pulex irritans, / förr du möjligen fanns. / Listig var du som räv och mård; / smög dig ända till kungens gård, / levde nästan i glans, / Pulex irritans."
Det finns två former av pest, böldpest med varbölder på huden och i lymfkörtlarna och lungpest som drabbar lungorna. Pesten finns numera inte i Sverige, men orsakar fortfarande tusentals dödsfall varje år i u-länderna. Dess spridning gynnas av dåliga hygieniska förhållanden, bland annat förekomst av råttor i nära kontakt med människor.

Pestloppan, Xenopsylla cheopis, är egentligen en råttloppa, men den är känd för att överföra den fruktade pestbakterien (Yersinia pestis) från svartråttor (Rattus rattus) till människor. Men numera står det klart, att pesten kan smitta människor på andra sätt än via svartråttornas loppor. Svartråttan saknades faktiskt i Nordeuropa under 1300-talet, då pesten härjade i form av digerdöden. Mellan människor kan pesten överföras av andra insekter än råttloppor, till exempel människoloppan, Pulex irritans, och troligen också av klädlusen, Pediculus humanus corporis, men inte av huvudlusen, Pediculus humanus capitis. Pesten kan också överföras direkt från människa till människa.
Digerdöden
Den mest beryktade pestepidemin är den som inleddes med digerdöden under medeltiden. Den hade sitt ursprung i Asien. Digerdöden nådde Europa omkring 1347 och Sverige via Norge 1350. Historiska data tyder på att mellan 30 och 50 procent av Europas befolkning dog inom loppet av fem år. Detta är så vitt vi vet den största katastrof som någonsin drabbat Europa i historisk tid. Dess hemska framfart beskrevs av den italienske författaren Giovanni Boccaccio i "Decamerone". Boccaccio omtalar att de drabbade ofta dog inom tre dagar efter det att de första symptomen hade uppträtt.
Denna epidemi återkom flera gånger fram till och med början av 1700-talet. De senare utbrotten blev inte lika allvarliga som digerdöden, delvis kanske beroende på att många av de människor som överlevde digerdöden hade en genetiskt betingad motståndskraft mot infektionen. Man införde också olika åtgärder för att förhindra pestens spridning. Till dem hörde karantän, av latinets "quadraginta", fyrtio, som syftar på det stipulerade antalet karantänsdagar. Fyrtio dagar var faktiskt ett korrekt val, eftersom pestens inkubationstid är cirka 37 dagar. England drabbades dock 1665–1666 av en stor pestepidemi, som beskrevs av Daniel Defoe, författaren till "Robinson Crusoe".
En del forskare har menat att digerdöden inte orsakades av pestbakterien utan av ett så kallat hemorragiskt virus, påminnande om det fruktade ebolaviruset. Men den teorin kan nu avskrivas. Forskarna har sekvenserat pestbakteriens hela arvsmassa. De har också isolerat DNA från pestbakterien ur resterna efter människor i medeltida pestkyrkogårdar. Pestbakterien härstammar från jordbakterien Yersinia pseudotuberculosis. Den förvärvade sin sjukdomsalstrande förmåga genom att ta upp två så kallade plasmider, som gjorde att den kunde infektera däggdjur. Plasmider är cirkelformade DNA-bitar, som inte ingår i bakteriens kromosom och kan överföras från en bakterie till en annan.
Andra pestepidemier
Vi känner till ytterligare två epidemier orsakade av pestbakterien. Båda hade troligen sitt ursprung i Asien. En började under 1800-talet och pågår fortfarande, numera dock i relativt liten skala. En annan pågick från och med 500-talet till och med 700-talet. Det mest berömda utbrottet är den så kallade justinianska pesten, som år 542 drabbade det östromerska rikets huvudstad Konstantinopel under kejsare Justinianus regeringstid. Denna epidemi kan ha varit en bidragande orsak till Östroms tillbakagång, tillsammans med två väldiga vulkanutbrott, som påverkade jordens klimat. Det står nu klart att epidemin orsakades av pestbakterien. Forskare har hittat bakteriens DNA i mänskliga kvarlämningar från pestepidemins tid.

De hittills äldsta fyndet av DNA från pestbakterien är från Sverige och har daterats till cirka 2 900 före vår tideräkning. Många stenåldersbosättningar försvann i Europa mellan 4 000 och 3 000 före vår tideräkning. Möjligen berodde det på en pestepidemi, men detta är inte klarlagt.
Det har också föreslagits att pesten fördes till Europa mellan 3 000 och 2 500 före vår tideräkning av människor från stäppen i öster. Vid denna tid invandrade nämligen till Europa ett mycket stor grupp människor tillhörande den så kallade Yamnayakulturen från området norr om Svarta havet. De använde så kallad snörkeramik och hade ceromoniella stridsyxor. Forskarna tror att att de till Europa införde det indoeuropeiska urspråket, från vilket nästan alla nutida europeiska språk härstammar. Möjligen underlättade pesten deras etablering i Europa, eftersom de kan ha haft större motståndskraft mot sjukdomen, än de människor som redan bodde i Europa. Arvet från dem upptar en stor del av nutida euopéers arvsmassa.
En annan svår epidemi var den så kallade pesten i Athen åren 430–426 före vår tideräknings början. Den beskrevs av Thukydides och drabbade athenarna hårt under det så kallade peloponnesiska kriget, då spartanerna hade invaderat deras land. Thukydides hävdar att spartanerna förgiftade athenarnas vattenreservoarer under belägringen av staden. Spartanerna kan alltså ha använt biologiska stridsmedel. Molekylärbiologiska undersökningar av daterat skelettmaterial har visat att pesten i Athen inte orsakades av pestbakterien, utan av bakterien Salmonella enterica serovar Typhi. Denna bakterie orsakar tyfoidfeber med inflammation av tunntarmens vägg, svår diarré och sepsis ("blodförgiftning"). Dödligheten är hög vid obehandlad tyfoidfeber.
Fördjupning
Huvudlöss, klädlöss, vårt förlorade kroppshår och de första kläderna. Du hittar artikeln under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
Referenser
- A. Andrades Valtueña et al.: The stone age plague and its persistence in Eurasia (Current Biology 27:3683-3691.e8, 2017).
- K. I. Bos et al.: A draft genome of Yersinia pestis from victims of the Black Death (Nature 478:506-510, 2011).
- E. Callaway: The Black Death decoded (Nature 478:444-446, 2011).
- M. Harbeck et al.: Yersinia pestis DNA from skeletal remains from the 6th century AD reveals insights into Justinianic Plague (PLoS Pathogens 9 [5]: e1003349, 2013).
- E. C. Holmes: Plague’s progress (Nature 478:465-466, 2011).
- A. K. Hufthammer och L. Walløe: Rats cannot have been intermediate hosts for Yersinia pestis during medieval plague epidemics in Northern Europe (Journal of Archaeological Science 40:1752-1759, 2013).
- M. J. Papagrigorakis et al.: DNA examination of ancient dental pulp incriminates typhoid fever as a probable cause of the Plague of Athens (International Journal of Infectious Diseases 10:206-214, 2006).
- M. J. Papagrigorakis et al.: The plague of Athens, an ancient act of bioterrorism? (Biosecurity and Bioterrorism: Biodefense Strategy, Practice, and Science 11:228-229, 2013).
- D. Raoult et al.: Plague. History and contemporary analysis (Journal of Infection 66:18-26, 2013).
- N. Rascovan et al.: Emergence and spread of basal lineages of Yersinia pestis; during the neolithic decline (Cell 176:295-305.e10, 2019).

Spökdjuren (några arter av släktet Tarsius) lever i den sydostasiatiska övärlden. De tillhör, tillsammans med halvapor och högre primater, ordningen primater bland däggdjuren. Till halvaporna hör bland andra lemurerna på Madagaskar och galagoerna i Afrika. De högre primaterna omfattar grupperna apor (östapor i Afrika och Asien samt västapor i Amerika) och hominoider (gibboner, orangutanger, gorillor, bonobon, schimpansen och människan). Det finns flera utdöda människoarter ("förmänniskor"), men alla nu levande människor tillhör samma art.
Spökdjuren intar något av en mellanställning mellan halvapor och högre primater. Spökdjurens välutvecklade synsinne och framåtriktade ögon är egenskaper som förenar dem med de högre primaterna. Spökdjurens ögon är enorma i förhållande till deras skalle. Om vi människor hade haft lika stora ögon till vår skalle, så skulle ögonen varit stora som äpplen. Stora ögon släpper in mera ljus, vilket gör dem ljuskänsligare, en fördel för nattaktiva djur. Spökdjurens ögon är så stora att de inte kan röra sig i ögonhålorna. I gengäld kan spökdjuren, i likhet med ugglorna, vrida huvudet 180 grader så att de tittar rakt bakåt.

Spökdjurens nos påminner också om nosen hos de högre primaterna. Den är nämligen kort och täckt med samma torra hud som finns i resten av ansiktet. Halvapornas nos är i regel längre och ytterst täckt med fuktig hud. Halvaporna har alltså precis som hunden en fuktig nos, ett så kallat rhinarium ("sniffbula").
Spökdjuren är nattaktiva rovdjur som lever i träden. Födan består av insekter och andra små djur. Spökdjuren har starkt förlängda bakben som gör dem mycket bra på att hoppa. Bakbenen är ungefär dubbelt så långa som kroppen. Märkligt nog är det vristens ben (i synnerhet hälbenet och båtbenet) som är mest förlängda.
Referenser
K. Gron och N. Grow: Primate factsheets. Tarsier (Tarsius), Taxonomy, morphology and ecology (Primate Info Net, University of Wisconsin-Madison, uppdaterad 21 november 2011).
P. Myers et al.: Tarsius, tarsiers (Animal Diversity Web, University of Michigan, hämtad 3 november 2016).

Komodovaranen (Varanus komodoensis) är den största nu levande ödlan. Det största uppmätta exemplaret lär ha varit 3,13 meter långt och vägt 166 kilogram. Vikten kan dock ha inkluderat en stor mängd ospjälkad föda i magtarmkanalen. En vikt på 70 kilo är mer normalt. Komodovaranen finns på den indonesiska ön Komodo och ett antal angränsande öar. Den lever till stor del på större djur, till exempel hjortar, som den sliter sönder med sina kraftiga sågkantade tänder.
Komodovaranen är en tandhygienists mardröm. I tändernas fördjupningar trivs nämligen en rikhaltig bakterieflora. Vi trodde förr att en komodovaran dödar sina bytesdjur genom att infektera dem med dessa bakterier och att bytena så småningom dör av "blodförgiftning" (sepsis). Men då måste varanen spåra upp bytet och risken är stor att en annan varan äter upp det.
Forskare har emellertid numera hittat gifter i saliven hos flera ödlearter, bland annat varaner och leguaner. Vi vet nu att komodovaranen har giftkörtlar, som mynnar i munhålan och att den dödar bytena med hjälp av gift, inte bakterier. Tidigare trodde forskarna att det bara fanns två giftiga ödlearter, gilaödlan ("gilamonstret") och dess släkting skorpiongiftödlan, båda i Nordamerika.

De största ödlorna vi känner till över huvud taget är de marina mosasaurierna som var toppredatorer i havet under yngre krita (cirka 100–65 miljoner år före nutid). Vissa mosasaurier kunde bli mer än 10 m långa. De var besläktade med dagens varanödlor.
Komodovaranens giftkörtlar mynnar i munhålan mellan tänderna. Tänderna är sågtandade med grunda fåror. De är inte försedda med djupa fåror eller giftkanaler, genom vilka giftet kan sprutas in i bytet. Djupa fåror finns bland annat hos de ovan nämnda arterna gilaödlan och skorpiongiftödlan, hos flera giftiga näbbmöss och hos många giftormar. Andra giftormar är försedda med kanaler i gifttänderna.
Komodovaranens gift innehåller flera komponenter. Två av dessa ämnen ger ett kraftigt blodtrycksfall, som troligen leder till cirkulatorisk chock, så att bytesdjuren inte kan fly. Ett annat ämne i giftet hämmar blodlevringen, så att bytet förblöder.
Komodovaranens skalle är inte särskilt kraftig och den kan inte bita med stor kraft. Sannolikt biter den tag i bytet och motstår dragkraften, när bytet försöker komma loss. Dess tänder åstadkommer då djupa sår. Därefter tömmer varanen giftkörtlarna i såren.
Hjärtat hos varanödlor är trerummigt, precis som hos andra ödlor. Men varanhjärtats kammare är försett med inbuktande skiljeväggar som gör att det fungerar nästan som däggdjurens fyrrummiga hjärta.
Fördjupning
Trerummare eller fyrarummare? Om fördelar med olika hjärtan. Du hittar artikeln under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
Referenser
Texten har uppdaterats och utökats år 2015.
- B.G. Fry et.al.: A central role for venom in predation by Varanus komodoensis (Komodo dragon) and the extinct giant Varanus (Megalania) priscus. (Proceedings of the National Academy of Science USA 106:8969-8974, 2009).
- Natural History Museum, London: Varanus prisca, giant monitor lizard (hämtad 18 februari 2015).
- Smithsonian Institution: Komodo dragon (hämtad 18 februari 2015).

Att kyssa en groda resulterar inte nödvändigtvis i att sagans prins plötsligt uppenbarar sig, utan du kan i stället få dig en mer oangenäm överraskning. De flesta grodor innehåller nämligen gifter. Dessa gifter ligger lagrade i hudkörtlar och utsöndras genom huden.
Att vara giftig är ett effektivt sätt att slippa undan rovdjur. Till världens giftigaste grodor hör pilgiftsgrodorna (familjen Dendrobatidae). Alla pilgiftsgrodor är inte lika giftiga och några är kanske inte giftiga alls. Det är farligt att bara vidröra de giftigaste arterna. Som syns på grodorna ovan och nedan har dessa grodor en färggrann teckning. Färgteckningen fungerar som en varningssignal (så kallad aposematism) för hugade spekulanter, som undviker att sluka grodan, när de ser färgerna. Det djur som haft turen att överleva, efter att ha slukat en pilgiftsgroda, glömmer inte färgteckningen. Detta utnyttjas av andra mindre giftiga grodor som har tagit efter de giftiga pilgiftsgrodornas färgteckning. Därigenom lurar de rovdjuren att tro att de är giftigare än vad de i själva verket är. Denna typ av falsk varubeteckning kallas mimikry.

Även människan har dragit nytta av pilgiftsgrodornas gift. Indianerna i vissa delar av Sydamerika använder grodornas gift till att smörja in sina pilspetsar med och gör därigenom pilarna till ett ännu dödligare vapen.
Pilgiftsgrodans gift består till största delen av kvävehaltiga föreningar som tillhör gruppen alkaloider. Alkaloiderna i giftet kan hos olika grodor variera både i antal och i grad av giftighet. En av de allra giftigaste alkaloiderna, batrakotoxin, leder vid förtäring till hjärtarytmier, andningsförlamning och hjärtstillestånd. Andra mindre giftiga alkaloider ger ökade muskelkontraktioner som kan övergå i muskelförlamning.
Pilgiftsgrodor som fötts upp i fångenskap, utan att få sin naturliga föda, har så småningom förlorat sina giftiga alkaloider. Forskare tror att grodorna får i sig de flesta, kanske alla, alkaloiderna genom att äta vissa insekter som finns i den naturliga miljön, till exempel myror. Sedan ackumulerar de alkaloiderna i hudkörtlarna. Insekterna får kanske i sin tur i sig alkaloiderna genom att äta vissa giftiga växter.
Även paddor (familjen Bufonidae) har giftkörtlar under huden och dessutom utsöndrar parotidkörtlarna (som är spottkörtlar) en del gift. Paddornas gifter skiljer sig kemiskt från pilgiftsgrodornas. Paddorna har en stor mångfald av olika ämnen i sina gifter. Paddornas gift utgör vanligen inte ett lika stort hot som pilgiftsgrodornas, men vissa paddor kan producera starka gifter. Agapaddan (Bufo marinus) och koloradoflodpaddan (Bufo alvarius) räknas till de giftigaste paddorna i världen och är kända för att ha förgiftat många katter och hundar. Intressant är, att en del rovdjur, till exempel igelkottar och illrar, är resistenta mot paddgifter.
Salamandrar utsöndrar också gift genom huden. Vissa salamandrar avger ämnet tetrodotoxin som är ett av de starkaste gifter djurvärlden har frambringat. Ett milligram tetrodotoxin är tillräckligt för att genom andningsförlamning döda en vuxen människa. Den färggranna europeiska eldsalamandern utsöndrar ett annat gift, salamandrin, i form av en mjölkaktig vätska från porer bakom ögonen. Detta sekret irriterar slemhinnor och ögon, och vållar smärtor för rovdjuren.
Referenser
J. W. Daly, H. . Garaffo, T. F. Spande, C. Jaramillo och A. S. Rand: Dietary source for skin alkaloids of poison frogs, Dendrobatidae? (Journal of Chemical Ecology 20:943-955, 1994)
A. Edström: Djurens gifter (Studentlitteratur, Lund, 1988)

En del blåsfiskar hör till de giftigaste fiskarna i världen. Trots, eller kanske tack vare, detta har en giftig blåsfisk kommit att bli betraktad som den yppersta av delikatesser i Japan. En måltid på denna fisk har liknats vid en gastronomisk version av rysk roulette.
Blåsfiskarna har fått sitt namn av att de snabbt kan fylla en blåsa i anslutning till matsmältningskanalen med vatten eller luft så att de från att normalt vara avlånga blir klotformiga. Förändringen sker vid fara som försvar mot angripare. Som en kompletterande försvarsstrategi har många blåsfiskar utvecklat en hög grad av giftighet. Blåsfiskfamiljen är vanlig i bl.a. Indiska oceanen och Stilla havet men förekommer inte i svenska vatten. Flera arter, särskilt från sötvatten och brackvatten, hålls som akvariefiskar.
Blåsfiskarnas gift heter tetrodotoxin eller TTX och är ett av de starkaste djurgifter vi känner. Det anses vara 25 gånger starkare än kurare och ett milligram är tillräckligt för att döda en människa. Giftet blockerar de spänningsreglerade kanaler för natriumjoner som finns i nervcellers och muskelcellers yttermembran. Strömmar av natriumjoner genom dessa kanaler är nödvändiga för att nervimpulser ska kunna uppkomma och förflytta sig. TTX-förgiftning leder därför till att den elektriska aktiviteten i nerver och muskler hämmas. I svåra fall leder detta till förlamning av andningsmusklerna med dödlig utgång.
Blåsfiskarna producerar inte giftet själva. Fiskar som kläckts och växt upp i fiskodlingar saknar nämligen TTX. Giftet produceras i stället av ett flertal marina bakteriearter. TTX-producerande bakterier har identifierats hos blåsfiskar, bland annat i mag-tarmkanalen och huden. Blåsfiskarna ackumulerar troligen TTX som produceras av dessa bakterier. Alternativt får fiskarna i sig giftet via bytesdjuren. I så fall härstammar giftet från bakterier som ätits av små djur på den lägsta nivån i näringskedjan och hamnar slutligen i blåsfiskarnas bytesdjur.
Vi kan fråga oss hur blåsfiskarna själva undviker att bli förgiftade. Forskare har emellertid hittat mutationer i de gener hos blåsfiskarna som kodar för proteinerna i natriumkanalerna. Mutationerna leder till att vissa aminosyror byts ut mot andra i proteinerna. Detta leder i sin tur till att blåsfiskarnas natriumkanaler inte påverkas av TTX.
TTX är inte unikt för blåsfiskar.

Blåsfisk kallas i Japan fugu och utgör grunden för en klassisk maträtt med samma namn. Den anses vara höjden av kulinarisk njutning och fisken betingar mycket höga priser. På grund av sin giftighet måste den tillagas av speciella, licensierade kockar. Särskilt levern, där giftkoncentrationen är högst, anses vara en gastronomisk upplevelse. Numera är det dock förbjudet att äta blåsfisklever. Ungefär 60 procent av alla fuguförgiftningar leder till döden och tidigare dog ett hundratal personer årligen i Japan på grund av sådana förgiftningar. På senare år har dock antalet dödsfall minskat.
I Japan används TTX i mycket utspädd form som ett smärtstillande medel vid bland annat reumatism. TTX har inom vetenskapen också visat sig vara ett oumbärligt hjälpmedel vid studier av nervceller och muskelceller på grund av sin förmåga att specifikt påverka natriumkanalerna.
Referenser
- E. G. Moczydlowski: The molecular mystique of tetrodotoxin (Toxicon 63:165-183, 2012).
- Nationalencyklopedin, band 3 (Bra Böcker, Höganäs, 1990).
- T. W. Soong och B. Venkatesh: Adaptive evolution of tetrodotoxin resistance in animals (Trends in Genetics 22:621-626, 2006).

Hur mycket tål du egentligen? Det spelar ingen roll hur mycket du har varit på gymmet eller hur tuff du känner dig, de små björndjuren tål mer!
Björndjuren, även kallade trögkrypare eller tardigrader, finns överallt på jorden där det finns fritt vatten någon gång på året, från Sibiriens tundra till oceanernas djup. De flesta lever i fuktig mossa. Björndjuren klarar temperaturer från åtminstone cirka -200 till cirka +150 grader Celsius. De klarar ren koldioxid, intensiv strålning och total uttorkning. Det är inte mycket som kan rå på björndjuren.
I ett fall lär en person ha tagit några totalt torra björndjur från en växt som hade befunnit sig i på ett museum i många decennier och fuktat dem lite och, minsann, en del av dem började röra på sig. Björndjur som hade legat infrusna i hela 30,5 år vid -20 grader Celsius i ett mossprov från Antarktis överlevde och fortplantade sig.
Trots att de kallas björndjur så är de inte så stora, som mest strax över en millimeter. De ser mycket märkliga ut med sina åtta ben, men är trots det inga spindlar. De påminner nästan mest om mycket förminskade däggdjur med lite väl många ben.
De små märkliga djuren kan vid kyla eller torka gå in i ett tillstånd som kallas för kryptobios. De ser då ut som små tunnor och slutar att förbruka syre. De skaffar sig inte energi på något annat sätt heller och vi skulle kunna tro att de var döda. Det handlar inte om liv på sparlåga, utan om ett tillstånd där livsprocesserna stannat av helt. Det är i detta kryptobiotiska tunnstadium som de kan överleva de extremaste förhållanden. Men så fort som förhållandena blir drägliga, till exempel om djuren tinar upp eller får lite fukt på sig, så lever de upp igen. Djuren dör inte ens när de är frusna i flytande kväve. Det är lite läskigt, det känns som en billig science fiction-film.
Hur klarar de då av kyla och torka? En viktig skyddsmekanism är att producera socker. Den innebär att björndjuren proppar cellerna och vätskan utanför cellerna fulla med sockerarten trehalos. Sockermolekylerna ersätter vatten och skyddar cellerna och deras membraner från uttorkningsskador. Även vid frysning finns det en stor risk för uttorkningsskador: om vatten binds i iskristaller uppkommer ju vattenbrist.

Det finns helt klart mycket forskning kvar att göra med björndjur och deras kryptobios, innan vi till fullo har förstått hur djuren kan tåla så extrema förhållanden.
Tillägg
En studie har visat att björndjur i kryptobiotiskt tillstånd klarar 6 000 atmosfärers tryck. Detta tryck är nästan sex gånger högre än trycket i oceanernas djupaste djuphavsgrav. Forskare har också identifierat ett protein som skyddar björndjuren mot röntgenstrålning.
Den svenske forskaren Ingemar Jönsson och medarbetare har visat att björndjur i kryptobiosis till och med klarar sig i rymden. Två arter av björndjur befann sig under tio dagar utanför en satellit som cirklade runt jorden. De hade inga problem med att överleva rymdens vakuum och den kosmiska strålningen. Däremot hade de problem med UV-strålningen från solen. Men ett fåtal djur överlevde även UV-strålningen. De ägg som dessa djur lade efter återkomsten till jorden kläcktes dock inte
De enda organismer som tidigare visat sig kunna överleva i rymden är endosporer av bakterien Bacillus subtilis och två arter av lavar. Endast ett mycket litet antal bakteriesporer överlevde UV-strålningen. Lavarna klarade däremot UV-strålningen alldeles utmärkt, förmodligen skyddade av det yttersta vävnadskiktet, barken. Lavar och bakteriesporer har även utsatts för krafter motsvarande dem som uppkommer när ett fragment av en planet slits loss vid en asteroidkollision och skjuts ut i rymden. Organismerna kunde överleva en sådan chock. De är således teoretiskt möjligt för levande organismer att transporteras mellan två planeter, till exempel Jorden och Mars, i synnerhet om de skyddas från UV-strålning genom att befinna sig inne i en meteorit. Vi vet att en del meteoriter, som fallit ner på jorden, härstammar från Mars.
Således är det fullt möjligt att livet uppkom på Mars och fördes till Jorden med meteoriter eller att det uppkom på Jorden och fördes till Mars. De flesta håller dock dessa alternativ för osannolika. Men vi får inte veta förrän vi upptäckt spår av liv på Mars, om det nu finns eller har funnits liv där.
Referenser
- G. Horneck et al: Microbial rock inhabitants survive hypervelocity impacts on Mars-like host planets: first phase of lithopanspermia experimentally tested (Astrobiology 8:17-44, 2008).
- K. I. Jönsson, E. Rabbow, R. O. Schill, M. Harms-Ringdahl, och P. Rettberg: Tardigrades survive exposure to space in low Earth orbit (Current Biology, 18: R729-R731, 2008).
- S. J. McInnes: Zoogeographic distribution of terrestrial/freshwater tardigrades from current literature (Journal of Natural History 28:257-352, 1994).
- K. Seki och M. Toyoshima: Preserving tardigrades under pressure (Nature 395:853-854, 1998).
- M. Tsujimoto, S. Imura, och H. Kanda: Recovery and reproduction of an Antarctic tardigrade retrieved from a moss sample frozen for over 30 years (Cryobiology 72:78-81, 2016).
- J. C. Wright, P. Westh och H. Ramløv: Cryptobiosis in Tardigrada (Biol. Rev. Cambridge Philosoph. Soc. 67:1-29, 1992).
Varför är vissa djur stora medan andra är små?

Varför har elefanter så oproportionerligt stora öron? Det har att göra med att elefanter är mycket stora djur som lever i ett mycket varmt klimat. Stora djur har svårt att göra sig av med den kroppsvärme som uppstår vid ämnesomsättningen och riskerar därför lätt att överhettas.
För att förstå elefanternas problem behöver vi titta på lite geometri. Om vi gör sidan på en kub dubbelt så lång, får vi en ny större kub.

En av den stora kubens ytor är fyra gånger större än en av den lillas ytor. Alltså är den stora kubens totala yta fyra gånger större än den lillas. Vi ser också att den större kuben rymmer åtta exemplar av den mindre. Alltså är den stora kubens volym åtta gånger större än den lillas. Vi förenklar det sedan lite och antar att en elefant är likadant formad som en mus och att elefanten är 100 gånger längre än musen (det vill säga elefanten är 5 m lång och musen 5 cm). Då blir elefantens kroppsyta 10 000 gånger (100 x 100) större än musens, medan dess volym blir hela 1 000 000 gånger (100 x 100 x 100) större än musens! Således har 1 deciliter elefant mycket mindre kroppsyta till sitt förfogande än 1 deciliter mus!
Både elefanten och musen producerar värme vid sin ämnesomsättning (metabolism), men elefanten och musen skulle omöjligt kunna producera lika mycket värme per deciliter djur. Båda djuren håller nämligen samma kroppstemperatur på cirka 36–38 °C. Vi vet faktiskt inte varför nästan alla däggdjur håller en kroppstemperatur på cirka 36–38 °C.

Musens problem blir då att den förlorar värme för lätt till omgivningen med sin stora yta per volymsenhet. Den måste därför ha en mycket intensiv ämnesomsättning, så att varje deciliter mus producerar tillräckligt med värme för att bibehålla hela musens kroppstemperatur.
Elefantens problem blir att den har svårt att göra sig av med värmen med sin lilla yta per volymsenhet. Den måste därför ha en lägre ämnesomsättning, så att den inte producerar mer värme per deciliter elefant än den kan göra sig av med. Gör den det kommer dess kroppstemperatur att stiga.
Om musen hade samma låga ämnesomsättning som elefanten, skulle den behöva isolera sig från värmeförluster med ett flera centimeter tjockt pälslager. Musen skulle inte kunna se något och den skulle inte kunna röra sig heller. Den skulle rulla runt i terrängen som en boll. Om elefanten hade samma höga ämnesomsättning som musen, skulle den inte hinna göra sig av med den värme den producerar. Den skulle då råka i kokning invärtes så att hela elefanten exploderade.

Yta-volymförhållandet räcker inte som förklaring
Förhållandet mellan yta och volym sätter alltså gränser för hur hög ämnesomsättning en elefant och en mus kan ha. Men det är mer komplicerat än så. Elefanten har faktiskt en högre ämnesomsättning i förhållande till musen än vi skulle förvänta oss, om det bara hade med yta-volymförhållandet att göra. Matematiskt uttryckt, så ökar ämnesomsättningen hos däggdjursarter mer när kroppsvikten ökar än förväntat, om ämnesomsättningen var direkt proportionell mot kroppsytan. Dessutom hittar vi samma samband mellan ämnesomsättning och kroppsvikt hos många djur som har samma temperatur i kroppen som i omgivningen. Så vi har ingen slutgiltig förklaring till detta märkliga samband som vi hittar inom väldigt många djurgrupper. Det finns flera hypoteser. En intressant hypotes hävdar att det har att göra med de trädlika förgreningarna hos bland annat blodkärlen. Dessa förgreningar följer de mönster som inom matematiken kallas fraktaler.
Hur elefanter svalkar sig i hetta
Men vilken funktion har då elefanternas öron? Jo, de stora öronen förbättrar deras yta-volymförhållande och ger dem en väsentligt större kroppsyta. Öronen är väl genomblödda och fungerar, med sin stora yta, som radiatorer genom vilka elefanten avger överskottsvärme från blodet till omgivningen. Flera andra anpassningar hjälper elefanter att avge värme. De har förlorat den värmeisolerande pälsen. Detta gör att de kan avge mer värme till omgivningen via värmeledning, så länge de är i skugga. Men de tar också upp mer värme i solsken. Den hårlösa huden underlättar emellertid värmeförluster genom avdunstning av kroppsvatten genom huden. Det åtgår nämligen värme, när flytande vatten övergår till gasform, alltså till vattenånga. Elefanter svettas inte, men kan ändå, tack vare hårlösheten, avdunsta mycket vatten genom huden. Dessutom är hudens yttersta döda hornlager inte jämnt. Det är försett med små upphöjningar, som är omgivna av fåror. Fårorna bildar ett bikakeliknande mönster. Det finns också en djupa veck i huden. När elefanter badar eller rullar sig i gyttja, fastnar vatten eller gyttja i hudens fåror och veck. Sedan avdunstar vatten från dessa håligheter. På så sätt kyls djuren utan att de förlorar vatten från kroppsvätskorna. Elefanter klarar sig dock inte utan att dricka vatten. De honor, matriarker, som leder hjordarna, vet sannolikt var det finns vattenhål och leder de törstande elefanterna dit.
Den tecknade elefanten Dumbos fysiologi
Den tecknade elefanten Dumbo torde ha större öron än någon annan elefant, åtminstone i relation till kroppsvikten. Nyligen har forskare genom datasimuleringar försökt utreda vad öronen betyder för Dumbos temperaturreglering. Först inhämtade de uppgifter om Dumbos dimensioner hos Disney. Forskarna kunde sedan uppskatta ämnesomsättningen hos en elefant i Dumbos viktklass. Simuleringarna antydde att Dumbo har svårt att hålla värmen när det är blåsigt och kallt. Öronen avger för mycket värme. Forskarna drog slutsatsen att Dumbo kanske behöver sina öron för att göra sig av med överskottsvärme när han flyger. Å andra sidan skulle Dumbo inte kunna flyga överhuvud taget, om han var en riktig elefant. Denna undersökning är inte bara ett skämt. Studier av påhittade djur kan ge information om vilka begränsningar som finns för riktiga djur.
Referenser
- X. Chen et al.: Growth, ageing and scaling laws of coronary arterial trees (Journal of the Royal Society Interface 12:20150830, 2015).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- G. Jayalalitha, V. Shanthoshini Deviha, och R. Uthayakumar: Fractal model for blood flow in cardiovascular system (Computers in Biology and Medicine 38:684-69, 2008).
- H. B. Lillywhite och B. R. Stein: Surface sculpturing and water retention of elephant skin (Journal of Zoology, London, 211:727-734, 1987).
- P.K. Phillips och J.E. Heath: Heat loss in Dumbo: a theoretical approach (Journal of Thermal Biology 26:117-120, 2001).
- K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).
- G. B. West, J. H. Brown, och B. J. Enquist: The fourth dimension of life: fractal geometry and allometric scaling of organisms (Science 284:1677-167, 1999).
- T. M. Williams: Heat transfer in elephants: thermal partitioning based on skin temperature profiles (Journal of Zoology, London, 222:235-245, 1990).
- P.C. Withers: Comparative Animal Physiology (Saunders College Publishing, 1992).

Varför är insekter små och däggdjur stora? De minsta nu levande däggdjuren är faktiskt ungefär lika stora som de största nu levande insekterna, i storleksordningen 5-15 cm långa. Vad begränsar däggdjurens och insekternas storlek?
Varför kan däggdjur inte bli hur små som helst?
Varför kan inte däggdjur bli hur små som helst? Det beror på att de alltid håller en högre kroppstemperatur än omgivningen, hos nästan alla äkta däggdjur (moderkaksdjur) cirka 36–38 °C. Mindre däggdjur har en större yta i förhållande till sin volym och avger därmed mer värme till omgivningen. Mindre däggdjur måste därför ha en högre ämnesomsättning, så att de producerar mer värme. Och för att hålla ämnesomsättningen hög måste de äta mycket mer, mätt per gram kroppsvikt, än större djur.
Små däggdjur, till exempel näbbmöss, måste alltså äta väldigt ofta och vi kan förstå att det bör finnas en nedre storleksgräns, under vilken det blir omöjligt att skaffa sig tillräckligt med föda. Små fåglar har samma problem som de små däggdjuren. Fåglarna har till och med större problem, eftersom som fåglar håller en så pass hög kroppstemperatur som cirka 39–41 °C. En del kolibrier skaffar sig en respit från det ständiga födosamlandet genom att under natten, då de inte kan samla föda, sänka kroppstemperaturen och gå i dvala. Sänkt kroppstemperatur innebär lägre ämnesomsättning. Detta gör att kolibrierna överlever natten på kroppens fettförråd, utan att behöva äta.
Varför är det bra att ha en hög kroppstemperatur?
Men varför måste, när de är vakna, äkta däggdjur vara cirka 36–38 grader och fåglar vara cirka 39–41 grader varma? Däggdjur och fåglar är jämnvarma vilket innebär att de håller en konstant kroppstemperatur som är högre än omgivningens. En fördel med detta är att det möjliggör en intensivare fysisk aktivitet och en större uthållighet vid löpning och flygning. En annan fördel är att djuren blir mindre beroende av vädret. De kan till exempel vara aktiva vid mycket låga yttertemperaturer. Det är ingen tillfällighet att det i Arktis finns däggdjur och fåglar, men inga växelvarma djur som grodor och reptiler. Men detta förklarar inte varför däggdjur håller sig just inom intervallet 36–38 grader och fåglar just inom intervallet 39–41 grader? Vi vet inte svaret på den frågan. Det finns mycket få äkta däggdjur och fåglar som håller lägre temperaturer än de angivna, trots att lägre temperaturer ger en lägre ämnesomsättning som i sin tur gör att djuren inte behöver äta lika mycket. En hypotes är att vatten eventuellt har vissa gynnsamma egenskaper inom det temperaturintervall som jämnvarma djur upprätthåller.
Varför kan insekter inte bli hur stora som helst?
Varför blir inte insekter större? Varför existerar inte sf-filmernas gigantiska insekter? För det första, så begränsas insekternas storlek kanske av deras yttre skelett (exoskelettet). Insekterna har inte skelettet inne i kroppen som vi, utan skelettet utgörs av det hårda kitinhaltiga lager (kutikulan) som utgör hudens yttersta skikt. Det finns mekaniska fördelar med ett exoskelett, men det har hävdats att ett exoskelett skulle ge problem för större djur. Ett problem skulle kunna vara hudömsningen (egentligen kutikulaömsningen). Tänk dig att vi människor, precis som insekter, under loppet av vår uppväxt skulle behöva genomgå ett antal kutikulaömsningar för att ge plats för ytterligare tillväxt. När vi förlorat den gamla kutikulan och den nya inte hunnit bildas, skulle vi helt sakna skelett. Denna övergångsfas borde ha varit rätt problematisk för ett stort landdjur som en människa. Vi skulle falla ihop som geléklumpar.
En ytterligare faktor som begränsar insekternas storlek är deras andningsapparat, trakésystemet. Detta system består av ett förgrenat träd av luftfyllda rör, vars finaste grenar ligger alldeles intill de enskilda cellerna. Transporten av syrgas i trakéerna sker, åtminstone i de finaste grenarna, med så kallad diffusion.

Mycket stora insekter skulle förmodligen få problem med alltför stora diffusionsavstånd. De finare trakégrenarna skulle bli för långa. Syrgastransporten skulle bli för långsam och vävnaderna skulle få för lite syrgas.
Men under karbonperioden (från cirka 359 till ca 299 miljoner år sedan) och permperioden (från cirka 299 till cirka 251 miljoner år sedan) fanns det trollsländor som var mycket större än någon nu levande insekt. De kunde ha en vingbredd på upp till cirka 6 decimeter! De levde emellertid under en tid då luftens syrgashalt var mycket högre än nu. De hade också en rätt smal kropp (cirka 3 centimeter), så diffusionsavstånden var inte så stora som du skulle kunna tro. Men det är nog ingen tillfällighet att det fanns andra jätteinsekter, till exempel kackerlackor, under den period då syrehalten i luften var hög. De blev dock inte så stora som sf-filmernas flugor. Vi hade kunnat brotta ner dem om vi stött på dem.
Den minskande syrehalten i luften från och med permperioden var sannolikt en mycket viktig bidragande orsak till att jätteinsekterna dog ut. Men jättetrollsländor fanns kvar, långt efter det att syrehalten hade blivit väsentligt lägre. Andra faktorer måste därför ha bidragit. Segelflygande och flygande dinosaurier kan till exempel ha konkurrerat ut dessa trollsländor eller haft dem som byte. Flygande dinosaurier kallas också fåglar.
Fördjupning
Hur sker transporter i djurens kroppar? Diffusion och massflöde. Du hittar artikeln under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
Referenser
Texten har uppdaterats och utökats år 2016.
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).
- UCMP web lift to taxa
- P.C. Withers: Comparative Animal Physiology (Saunders College Publishing, 1992).

Hur stort kan ett flygande djur bli utan att förlora förmågan att flyga? Vilken är det största flygande fågel vi känner till? Hur stor kan en fågel som förlorat flygförmågan bli?
Mycket stora fåglar förlorar flygförmågan
De största fåglar vi känner till har alla förlorat flygförmågan. Den tyngsta nu levande fågeln är den afrikanska strutsen (Struthio camelus). De största kända utdöda fåglarna troddes intill nyligen ha varit elefantfågeln Aepyornis maximus på Madagaskar, som sannolikt var besläktad med strutsfåglarna, och den australiska Dromornis stirtoni som troligen härstammade från andfåglarna. Forskare anser att dessa fåglar kunde väga upp till cirka 500 kg. År 2018 beskrevs emellertid en ännu större utdöd elefantfågel (Vorombe titan) vars vikt, baserat på skelettbenens storlek, uppskattats till i medeltal cirka 640 kg. Alla dessa fåglar härstammade från fåglar med flygförmåga. Alla nutida icke flygkunniga fåglar härstammar också från flygande fåglar.
Numera är forskarna tämligen ense om att fåglarna härstammar från små teropoder. Teropoderna tillhör dinosaurierna och därmed är fåglarna inte en giltig systematisk grupp. De är teropoder och därmed reptiler. De största teropoderna var Tyrannosaurus rex och några av dess släktingar vars vikter uppskattats ha legat mellan 10 och 20 ton. Uppskattningarna är osäkra och det är oklart vilken av dem som var tyngst. Det har spekulerats att det skulle finnas en övre gräns på cirka 12–15 ton för deras kroppsvikt, men det är oklart vilken faktor som i så fall skulle vara begränsande.

Jag har svårt att hitta några fysiologiska orsaker som skulle hindra icke flygande fåglar att nå samma storlek som de största teropoderna. Utdöda teropoder gick ju på två ben precis som fåglar och var inte så olika dagens strutsfåglar. Mot denna tanke talar dock det faktum att vi inte känner till några fåglar i denna viktklass.

De största kända flygande fåglarna
När det gäller flygande fåglar finns en övre gräns för kroppsvikten. Svanar, kondorer och trappar tillhör de tyngsta nu levande flygande fåglarna. Den tyngsta av dem alla anses vara den afrikanska koritrappen (Ardeotis kori), med en vikt på cirka 16 kg eller mer. Den anses kunna bli tyngre än den andinska kondoren, som ofta nämns i sammanhanget. Den utdöda argentinska kondorliknande Argentavis magnificens uppskattas vara den tyngsta kända flygande fågeln, med en uppskattad vikt på upp emot cirka 80 kg, men uppskattningen är mycket osäker. Dess likaledes utdöda släkting Pelagornis sandersi uppskattas ha haft en vikt på cirka 22–40 kg, men även denna uppskattning är osäker. Pelagornis uppges ha haft den största kända vingspannet bland fåglarna, 6,4 meter, dubbelt så brett som hos en albatross.

De största flygande djuren någonsin var flygödlor
Vissa arter bland de utdöda flygödlorna (egentligen bör de kallas pterosaurier, de var varken ödlor eller dinosaurier) var också mycket tunga. Dessa arter kunde troligen flyga. Den största av dem alla var Quetzalcoatlus northropi. Uppskattningarna av dess vikt varierar från cirka 60–70 kg till cirka 200–250 kg. Den är troligen det största kända flygande djuret. Dess vingspann har uppskattats till cirka 10 meter. Flygödlorna flög med hjälp av en flyghud, precis som fladdermöss.
Hatzegopteryx thambema från Rumänien var kanske större än Quetzalcoatlus. Men av den har forskarna bara hittat ett fåtal skelettben. Dett är oklart hur stor den var och om den kunde flyga.

Hur kunde de största flygande djuren flyga?
För att en fågel ska flyga måste den kunna utveckla en tillräckligt stor effekt med sina muskler. Effekten mäts i watt eller hästkrafter, precis som för en bilmotor. Fåglarna måste alltså ha en tillräcklig effektiv "motor". Ju större en fågel är, ju mindre effekt kan den utveckla per kilo kroppsvikt. Stora fåglar är alltså missgynnade och det måste finnas en gräns för kroppsvikten, över vilken flygande blir omöjligt. Men teoretiska beräkningar visar att de utdöda stora fåglarna och flygödlorna mycket väl kan ha utvecklat tillräckligt stor effekt. Stöd för denna tanke ger flygplanet Gossamer Albatross som 1979 flög över Engelska kanalen drivet av en tävlingscyklist.
De allra största flygande djuren kunde förmodligen inte flyga längre sträckor aktivt med flaxande vingar. Men de var troligen väl anpassade till att glidflyga med hjälp av uppvindar. Detta är betydligt mindre energikrävande än aktiv flygning. Uppvindar förekommer på slät mark, då uppvärmd luft stiger uppåt (termik). De uppkommer också då vindar möter berg och höjder i terrängen eller vågor på havet. Många av de största nutida flygande fåglarna, till exempel gamar, albatrosser och kondorer är experter på att utnyttja uppvindar. Gamar kan glidflygande stiga till flera tusen meters höjd med hjälp av termik. Även människor kan använda uppvindar för att glidflyga långa sträckor, dock med hjälp av segelflygplan, hängglidare eller fallskärmar.
Stora flygande fåglar har emellertid svårt att lyfta från marken. Vid vindstilla måste de, precis som flygplan, nå en viss hastighet för att av fartvinden få så stor lyftkraft att de släpper markkontakten. Svanar springer ofta långa sträckor på vattenytan och albatrosser på marken, innan de når tillräcklig hastighet för att bli luftburna. Pterosauriernas bakben låg inuti flyghuden och bidrog till att spänna ut den vid flygning. Detta innebär att de knappast kunde springa särskilt fort. I en studie beräknades att en Quetzalcoatlus på 75 kg skulle kunna lyfta i måttlig motvind springande med bakbenen, medan en Quetzalcoatlus på 250 kg skulle ha behövt en orimligt kraftig motvind för att kunna lyfta. Motvinden adderas nämligen till fartvinden och ökar lyftkraften. Det är välkänt att svanar och albatrosser föredrar att lyfta i motvind. Vid stark motvind kan det, även för stora djur, i princip räcka med att de breder ut vingarna för att de ska kunna lyfta. Men det vore en stor nackdel att vara helt beroende av vinden för att lyfta och bli helt markbunden i stiltje. Ett alternativ är att tillbringa den icke flygande tiden på klipphyllor och eller andra högt belägna ställen, ungefär som tornseglare, och bara kasta sig ut i luften för att bli luftburen. Tornseglare är mycket små fåglar, men har ändå mycket svårt att lyfta från marken. Forskare har framkastat tanken att många jättestora flygande djur levde ett liknande liv och aldrig landade på slät mark.

Gjorde jätteflygödlorna avstamp mot marken med både bakben och vingar?
Ovanstående resonemang bygger på att de utdöda jättestora flygande djuren bara använde bakbenen för att lyfta från marken. Detta skulle kunna gälla för fåglarna, men inte för flygödlorna. Deras vingar var helt annorlunda uppbyggda än fåglarnas. Detta gällde både muskulatur och skelett. Dessutom flög de med hjälp av flygmembran, inte med hjälp av fjädrar. Vingmembranet (flyghuden) stöddes av ett kort överarmsben (humerus), långa underarmsben (radius och ulna), långa mellanhandsben (metacarpalia) samt ett oerhört långt fjärde finger med fyra fingerben (falanger). Handlovsbenen var reducerade. Vid handloven fanns dock ett för flygödlorna unikt ben, (pteroideus), mer om det senare. Tre mycket små fingrar fanns vid det fjärde fingrets bas. Se bilden nedan.
Forskarna tror att flygödlornas skelettben i mycket hög utsträckning var luftfyllda, vilket minskade deras vikt. Men, precis som hos fåglar, var benen förstärkta av tvärgående balkar, något som gav dem en hög hållfasthet. Detta kan ha varit nödvändigt för att de skulle kunna flyga över huvud taget. Vingarnas uppbyggnad gjorde att flygödlorna sannolikt kunde stå och gå på fyra ben, precis som fladdermöss. De vilade då på bakfötterna och på det fjärde fingrets bas vid leden mellan fingret och mellanhandsbenen i vingarna. När den största kända flygödlan, Quetzalcoatlus northropi stod på marken, var den lika hög som en giraff.

I en omfattande studie från 2014 tog forskarna fasta på att flygödlornas unika anatomi. De räknade med att Quetzalcoatlus northropi vägde 180–250 kg, hade ett vingspann på 10 m och upp till 50 kg flygmuskler. Forskarna lanserade en helt ny idé som skulle kunna göra de tyngsta flygödlorna kapabla att lyfta direkt från marken. Om de kunde stå på bakbenen och vingarna, borde de kunna utnyttja inte bara bakbenen utan också vingarna med sina flygmuskler till att lyfta och direkt bli luftburna. Enligt denna modell hukade de sig ner genom att böja knäleden och vingarnas leder. Sedan rättade de ut bakbenen, vilket gjorde att de roterade framåt. Slutligen rätade de ut vingarna med hjälp av de massiva flygmusklerna. Detta resulterade i att de hoppade högt upp i luften, som skjutna ur en katapult, vecklade ut vingarna och omedelbart blev luftburna. Kanske spändes senor i vingarna och lagrade elastisk lägesenergi, som i en spänd pilbåge. Denna lägesenergi kunde vid avstampet omvandlas till rörelseenergi och, jämte muskelsammandragningarna, bidra till att flygödlan lyfte från marken. De elastiska senorna kunde kanske låsas i spänt läge och utlösas med en avtryckarmekanism.
Forskare har testat modeller av flygödlor i vindtunnlar. Flygödlornas flyghud bestod av flera delar. Huvuddelen av flyghuden var spänd mellan vingens ben och bakbenen. Det fanns också flyghud bakom bakbenet och, i form av en så kallad framvinge, framför vingens ben. Den sistnämnda flyghuden spändes ut av ett skelettben (pteroideus) som var fäst i handloven och bara hittats hos flygödlor, se bild högre upp på sidan. Detta ben hade troligen stor betydelse för flygödlornas flygförmåga. Med benet riktat framåt spändes modellernas framvinge ut och gav hela vingen förbättrade aerodynamiska egenskaper. Kanske kunde framvingen också vinklas i olika lägen, uppåt och neråt.
För att summera så kan det trots allt vara svårigheten att lyfta som sätter en övre gräns för kroppsvikten hos flygande djur, men att djur, som var mycket tyngre än nutidens tyngsta fåglar, sannolikt kunde vara utmärkta flygare.
Fördjupning
Hur började fåglarna flyga? Om tvåbenta dinosaurier, fjädrar och vingar. Du hittar artikeln under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?.
Vilka djur hoppar högst? Hur kan små djur, som loppor och gräshoppor, hoppa så högt? Artikeln finns under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?.
Referenser
- R. M. Alexander: All-time giants: the largest animals and their problems (Palaeontology 41:1231–1245, 1998).
- E. Buffetaut, D. Grigorescu, och Z. Csiki: A new giant pterosaur with a robust skull from the latest Cretaceous of Romania (Naturwissenschaften 89:180-184, 2002).
- M. Habib: Anatomy (Pterosaur.net, hämtad 30 september 2016).
- J. P. Hansford och S. T. Turvey: Unexpected diversity within the extinct elephant birds (Aves: Aepyornithidae) and a new identity for the world's largest bird (Royal Society Open Science 5:181295, 2018).
- D. T. Ksepka: Flight performance of the largest volant bird (Proceedings of the National Academy of Sciences 111:10624–10629, 2014).
- C. Palmer: Flight in slow motion: aerodynamics of the pterosaur wing (Proceedings of the Royal Society B 278:1881-1885, 2011).
- P. Palmqvist och S. F. Vizcaíno: Ecological and reproductive constraints of body size in the gigantic Argentavis magnificens (Aves, Theratornithidae) from the Miocene of Argentina (Ameghiniana 40:379-385, 2003).
- F. Therrien och D. M. Henderson: My theropod is bigger than yours ... or not: estimating body size from skull length in theropods (Journal of Vertebrate Paleontology 27:108-115, 2007).
- University of Portsmouth: Prehistoric winged reptiles 'pole-vaulted' into flight (ScienceDaily, 15 November 2010).
- M. T. Wilkinson, D. M. Unwin och C. P. Ellington: High lift function of the pteroid bone and forewing of pterosaurs (Proceedings of the Royal Society B 273:119–126, 2006).
- M. P. Witton och M. B. Habib: On the size and flight diversity of giant pterosaurs, the use of birds as pterosaur analogues and comments on pterosaur flightlessness (PLoS ONE 5[11]: e13982).
- M. P. Witton och D. Naish: A reappraisal of azhdarchid pterosaur functional morphology and paleoecology (PLoS ONE 3[5]: e2271).
Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?

Djurens kroppsform har stor betydelse för deras funktioner och överlevnad. De mest grundläggande skillnaderna mellan olika djurgrupper gäller kroppens symmetri. Tre av de största djursstammarna uppvisar så kallad radiärsymmetri, nästan alla andra så kallad bilateralsymmetri. De senare brukar kallas tvåsidiga djur. Varför är de flesta djur, även människor, bilateralsymmetriska med högersida och vänstersida?
Om ett plant snitt genom ett djur ger upphov till två kroppshalvor, som är spegelbilder av varandra, kallas snittet ett symmetriplan (spegelplan). De enklast uppbyggda djurstammarna, svampdjuren och de så kallade plakozoerna, saknar symmetriplan. De är oregelbundet uppbyggda. Alla andra djur har symmetriplan. Det förekommer dock ofta större eller mindre avvikelser från symmetrin, särskilt inuti kroppen. Det finns två huvudtyper av symmetri: radiärsymmetri (radialsymmetri) och bilateralsymmetri. Biradiärsymmetri är ett specialfall av radiärsymmetri. Tvåsidiga djur är djur med bilateralsymmetri. Denna term är inte lyckad. Alla djur har ju flera sidor.

Radiärsymmetri
Några djurstammar uppvisar radiärsymmetri. Det innebär att vi kan skära flera olika plana snitt genom djurets kropp, vilka alla ger upphov till två kroppshalvor som är spegelbilder av varandra. dessa djur har således flera symmetriplan (spegelplan). De flesta sjöstjärnor är femarmade med munnen på undersidan. De har femtalig radiärsymmetri. Från översidan mot undersidan kan du genom sjöstjärnan skära fem olika snitt, som bildar spegelplan. Varje snitt går genom en armspets och genom spetsen av en av de vinklar som bildas mellan armarna. Hjul är radiärsymmetriska. Ett hjul med fem symmetriskt placerade ekrar har femtalig radiärsymmetri, om utsidan av hjulet, svarande mot djurets översida, ser annorlunda ut än insidan, svarande mot djurets undersida med dess mun.

Nässeldjur (till exempel maneter och havsanemoner) är ursprungligt radiärsymmetriska. De placeras nära basen av djurens släktträd. De kan ha olika antal symmetriplan, bland annat beroende på hur tentakler, sinnesorgan och inre skiljeväggar är placerade. Vissa av dem är biradiärsymmetriska, åtminstone i sin inre uppbyggnad.
Vuxna tagghudingar (till exempel sjöstjärnor och sjöborrar) är också radiärsymmetriska, dock med vissa avvikelser från symmetrin. De härstammar emellertid från bilateralsymmetriska djur (bilateralsymmetri förklaras nedan). Detta frangår bland annat av att deras larver är bilateralsymmetriska. Märkligt nog ligger tagghudingarna nära ryggradsdjuren på djurens släktträd.
Tagghudingar är kända ända sedan den kambriska explosionen för cirka 500 miljoner år sedan, då de flesta av dagens större djurstammar uppträdde. De har överlevt alla de katastrofiska utdöenden, som ägt rum sedan dess, och är än i dag en artrik och framgångsrik djurgrupp. Fördelen med deras övergång från bilateralsymmetri till radiärsymmetri är en gåta. De flesta av dem lever nämligen på havsbottnen, där deras kroppssymmetri sannolikt gör dem mindre rörliga. Deras uppbyggnad är så märklig, att vi skulle kunna tro att de kom från en annan planet.
Du skulle kunna tro att bläckfiskarna är radiärsymmetriska. De åttaramade bläckfiskarnas armar uppvisar i och för sig radiärsymmetri. Men resten av kroppen är bilateralsymmetrisk hos alla bläckfiskar. Tioarmade bläckfiskar har åtta kortare armar och två längre tentakler, vilket gör hela kroppen bilateralsymmetrisk. Du skulle också kunna tro att daggmaskar, med sin cylindriska kroppsform, är radiärsymmetriska. Men det är de inte, Deras inre uppbyggnad är tydligt bilateralsymmetrisk.
Biradiärsymmetri
Ett specialfall av radiärsymmetri är biradiärsymmetri. Det innebär att det finns två symmetriplan (spegelplan). Ovannämnda hjul blir biradiärsymmetriskt, om du förser det med endast två motställda ekrar.

Kammaneterna är den enda djurstammen med uteslutande biradiärsymmetrisk byggnad. De utgör en enkelt byggd djurstam, dock mer komplext uppbyggda än nässeldjuren. De brukar placeras tillsammans med nässeldjuren, nära basen av djurens släktträd. Nya molekylära data tyder dock på att kammaneterna är en systergrupp till alla andra djur, inklusive svampdjuren. Fantastiskt nog, skulle detta innebära att muskelceller och nervceller hos djur utvecklats två gånger, oberoende av varandra. Men detta är fortfarande mycket kontroversiellt.
Bilateralsymmetri
Vi däggdjur och de flesta andra djur har bara ett enda symmetriplan. Planet delar oss i en högersida och en vänstersida, vilka är spegelbilder av varandra. Detta kallas bilateralsymmetri. Bilateralsymmetriska djur har en buksida (ventralsida) och en ryggsida (dorsalsida) som är olika, samt en framände (kranial eller rostral ände) och en bakände (kaudal ände) som också är olika.

Bilateralsymmetri är en evolutionär anpassning till att lätt kunna röra sig. En särskilt utformad framände med sinnesorgan underlättar rörelsen framåt. Människor rör sig dock inte med framänden först utan med ventralsidan, såvida de inte simmar eller kryper.
Olika utformning på buksidan och ryggsidan får ses som en evolutionär anpassning till tyngdkraften, som alla djur utsätts för. Tack vare denna anpassning kan simmande och flygande djur röra sig lättare och hålla balansen. Djur som rör sig på ett fast underlag kan röra sig med hjälp av en krypsula på buken. Avancerat uppbyggda bilateralsymmetriska djur har på buken i stället extremiteter, som gör att de rör sig snabbare och lättare. Exempel är leddjurens och fyrfotadjurens gångben.
Grävande ryggradlösa djur är ofta cylindriska med en till det yttre mindre tydlig bilateralsymmetri, men med bilateralsymmetriska inre organ. Detta beror säkerligen på att tyngdkraften har mindre betydelse nere i jord eller bottensediment. I trånga gångar skulle det det till med kunna vara en fördel att sakna tydligt utvecklad ryggsida och buksida.
Bilateralsymmetri har hos de flesta frilevande djurgrupper lett till att en hjärna, flera olika sinnesorgan samt munnen lokaliserats till framänden i ett huvud. Detta är naturligtvis funktionellt, eftersom det är framänden som först kommer i kontakt med sinnesintryck och föda.
Hos bilateralsymmetriska djur finns det ofta vissa avvikelser från symmetrin kring det enda symmetriplanet, i synnerhet avseende de inre organen. Vi däggdjur har ju till exempel levern till höger, mjälten och magsäcken till vänster samt hjärtat förskjutet mot vänster sida.
Finns det komplicerat byggda flercelliga organismer på himlakroppar utanför vårt solsystem? Vi vet inte. Själv tror jag att universum är så stort att det vore mycket konstigt, om sådana inte fanns. Och jag är övertygad om att de flesta av dem har bilateral symmetri, om de kan röra på sig. De lever trots allt, precis som vi, på en tredimensionell planet eller måne med tyngdkraft. Fysikens lagar gäller i hela universum.
Fördjupning
Vad är djur? Vad skiljer dem från andra organismer? Hur uppkom djuren? Du hittar artikeln under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd.
Hur nya ben och svansar ersätter gamla. Regeneration: återväxt av amputerade kroppsdelar. Artikeln finns under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?.
Referenser
- R.C. Brusca och G.J. Brusca: Invertebrates (2nd ed, Sinauer, 2003).
- C. Hickman, L. Roberts, A. Larson, H. l'Anson, och D. J. Eisenhour: Integrated principles of zoology (13th ed,, McGraw Hill, 2005).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- L.L. Moroz et al.: The ctenophore genome and the evolutionary origins of neural systems (Nature 510:109-114, 2014).
- P. Simion et al.: A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals (Current Biology 27:1-10, 2017).

Med skelett avses ofta stödjande delar av djurkroppar som består av hårt material, antingen ett yttre skelett (exoskelett) eller ett inre (endoskelett). Det finns emellertid många djur som saknar ett hårt skelett. Exempel är maneter, plattmaskar och daggmaskar. Små djur klarar sig bättre än stora utan att ha ett hårt skelett. Många små djur saknar därför skelett. Andra djur, både små och stora, har ett mjukt skelett som kallas vätskeskelett eller hydrostatiskt skelett.
Vattenlevande djur klarar sig bättre är landlevande utan att ha ett hårt skelett, eftersom vattnet bär upp kroppen. I vatten kan därför mycket stora djur, som de flesta bläckfiskar, klara sig utan hårt skelett. Grävande djur klarar sig också lättare utan hårt skelett, eftersom den omgivande jorden ger stöd. Det finns daggmaskar som är ett par meter långa.

Vätskeskelett (hydrostatiskt skelett)
Men de flesta av de djur som saknar hårt skelett får stadga i kroppen på annat sätt och de kan därför sägas ha någon form av mjukt skelett. Oftast handlar det om en sluten vätskefylld kroppshåla, ett så kallat hydrostatiskt skelett (vätskeskelett). Längsgående och ringformade muskler finns då runt hålan. Musklerna verkar på kroppshålan, så att den ändrar form. En vätska kan ju inte pressas ihop så att dess volym minskar. När de ringformade musklerna dras ihop, blir därför kroppen längre och smalare. När de längsgående musklerna dras ihop, blir den kortare och tjockare. När de längsgående musklerna på en sida av kroppen dras ihop, böjs den så att dess framände och dess bakände vänds åt denna sida. Ett hydrostatiskt skelett fungerar bättre om det är uppdelat i flera separata håligheter. Så är det ordnat hos daggmaskar och havsborstmaskar, vars kroppar är uppdelade i ett stort antal segment. Rundmaskar (nematoder) har inga ringformade muskler, bara längsgående. Deras hydrostatiska skelett och deras yttre skelett (kutikulan) ger upphov till motkrafter som rätar ut masken, när de längsgående musklerna på en sida av kroppen slappnar av.

Muskler kan fungera som skelett
Ett mjukt skelett kan också vara en så kallad muskulär hydrostat, som bara består av muskler. I en sådan fungerar musklerna som ett icke sammanpressbart hydrostatiskt skelett, samtidigt som de åstadkommer rörelse. Bläckfiskarnas armar, elefantens snabel och människans tunga är alla muskulära hydrostater.

Inre och yttre hårda skelett (endoskelett och exoskelett). Hur muskler verkar på dem
Hårda skelett finns av två typer, inre skelett (endoskelett) och yttre skelett (exoskelett). Alla ryggradsdjur (människor och andra däggdjur, kräldjur inklusive fåglar, groddjur och fiskar) har ett inre skelett som ligger inne i kroppen och påverkas av muskler som ligger utanför skelettet. Leddjur (till exempel kräftdjur, spindlar och insekter) har ett yttre skelett med muskler som arbetar innanför skelettet. I båda fallen är en muskel fäst på båda sidorna av en skelettled. Leden fungerar som ett gångjärn. Avstånden mellan muskelns fästpunkter och leden kan vara olika stora. Vi får då olika typer av hävstångseffekter. Sådana effekter kan bland annat leda till att muskelkraft förloras, men att rörelsens spännvidd och hastighet i stället blir större.
De flesta blötdjur (till exempel musslor och snäckor) har ett yttre skelett i form av ett skal. Tagghudingar (till exempel sjöstjärnor och sjöborrar) har faktiskt ett inre skelett. En sjöstjärna är hård att ta på, men skelettet ligger innanför hudens yttersta skikt och är därför ett inre skelett.
Ryggradsdjurens inre skelett: ben och brosk
Skelett innehåller i regel många olika kemiska ämnen. Benen i vårt och de flesta andra ryggradsdjurs skelett blir hårda av hydroxiapatit. Detta ämne är ett mineral och består av kristaller, som huvudsakligen innehåller kalciumjoner och fosfatjoner. Skelettbenen blir sega med hjälp av trådformiga proteiner, så kallade kollagener, som motstår sträckning och böjning. Hydroxiapatiten och kollagenerna bildar en så kallad matrix utanför benvävnadens celler. Benvävnaden blir extra stark på grund av att den innehåller två komponenter med olika egenskaper, precis som armerad betong. Järnstavarna i betongen fungerar som kollagenet, betongen som hydroxiapatiten.
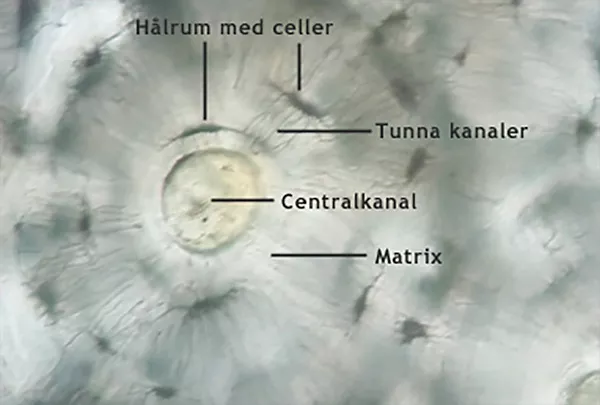
Hajar och en del andra fiskar saknar benvävnad i skelettet, utom tänderna, och har i stället bara brosk. Brosk ingår också i andra ryggradsdjurs skelett. Brosk finns hos människan bland annat i struphuvudet, ytteröronen och nässpetsen samt på ledernas ytor. Brosk innehåller utanför cellerna en matrix bestående av ett nätverk av kollagenfibrer omgivet av så kallade proteoglykaner. De senare utgörs av stora komplex innehållande långa kolhydratkedjor, bland annat kondroitinsulfat och hyaluronsyra, och proteiner. Brosk är mer eller mindre böjligt och ibland elastiskt, men inte lika hållfast som benvävnad.

Ryggradslösa djurs yttre och inre skelett
Insekters och andra leddjurs yttre skelett kallas kutikula. Kutikulan bildas av hudens yttersta cellskikt. Den innehåller kedjeformade molekyler av kolhydraten kitin och ett flertal olika proteiner. Proteinmolekylerna är bundna till varandra, så att de bildar ett nätverk. Kräftdjurens skelett blir dessutom extra hårt av kalciumkarbonat, ett ämne som även finns som mineraler i kalksten. Insekter och andra leddjur måste då och då kasta av det gamla yttre skelettet och anlägga ett nytt större skelett. Annars skulle skelettet hindra dem från att växa och bli större. Vi säger att de "ömsar hud" eller att de "ömsar skal".
Blötdjurens skal innehåller huvudsakligen kalciumkarbonat och sammanbundna proteiner som kallas konkiner. Skalet bildas av den del av huden som kallas manteln. Blötdjur byter inte skal. Skalet växer i kanterna, när djuret växer.
Tagghudingarnas skelett bildas under det yttersta hudskiktet och innehåller sammanbundna plattor eller spridda bitar av kalciumkarbonat. Inuti kalciumkarbonatet finns ett nätverk av kanaler, som innehåller celler och fibrer.
De mycket enkelt uppbyggda svampdjuren har faktiskt ett inre skelett. De flesta svampdjur har ett hårt skelett som innehåller kalciumkarbonat eller kiseldioxid. Flera mineraler, till exempel kvarts, består också av kiseldioxid.

Riktigt stora landdjur som människor och de flesta andra däggdjur hade förmodligen inte klarat sig med ett yttre skelett, som måste ömsas för att tillåta tillväxt. Ömsning fungerar bra för små djur som har rätt mycket stadga även när de kastat av sig det gamla skelettet och inte hunnit bilda ett nytt. Men om människobarnen hade gjort på det viset hade de sjunkit ihop som geléklumpar när de kastat av det gamla skelettet och det hade varit omöjligt för dem att gå till skolan.
Fördjupning
Hur kryper daggmaskar och plattmaskar? Du hittar artikeln under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur? här nedanför.
Varför är insekter så små? Varför är däggdjur och fåglar stora? Artikeln finns under rubriken Varför är vissa djur stora medan andra är små?
Referenser
- R. C. Brusca and G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- A. Stevens och J. Lowe: Human histology (3rd ed, Elsevier Mosby, 2005).
- G. J. Tortora och M. T. Nielsen: Principles of human anatomy (12th ed, Wiley, 2012).
Människor och andra djur rör sig genom att muskler dras ihop. En muskel med sin sena påminde tidiga anatomer om en mus med svans och kallades därför "musculus" ("liten mus" på latin). Först trodde många att muskler förkortas genom att proteinmolekyler drar ihop sig inuti musklernas celler. Men kring mitten av 1900-talet kom forskare underfund med att de aktiva muskelproteinerna inte förkortas, utan glider mot varandra, den så kallade filamentförskjutningsmekanismen.
I det följande förklaras först muskelcellens uppbyggnad, sedan hur en skelettmuskel drar ihop sig, kontraheras.
Muskelcellens uppbyggnad
Bilden nedan visar en sarkomer, den grundenhet som ingår i skelettmusklernas celler. Skelettmuskelcellerna utgörs av mycket långa och tjocka muskelfibrer. Inuti muskelfibrerna är sarkomerer ihopkopplade efter varandra i långa rader. Många sådana rader bildar en myofibrill. Varje muskelfiber innehåller ett stort antal parallella myofibriller.

Alla sarkomerens delar består av proteiner. Sarkomeren avgränsas på båda sidorna av tvärgående Z-linjer. De ingår också i angränsande sarkomerer, hopkopplade med den ovan avbildade. Flera långsträckta tunna aktinfilament är fästa i Z-linjerna, varje filament bestående av en mängd sfäriska aktinmolekyler. Mellan aktinfilamenten finns tjocka myosinfilament, varje filament bestående av ett stort antal myosinmolekyler. Den tvärgående M-linjen kopplar ihop myosinfilamenten. Varje myosinmolekyl är i sin ena ände försedd med ett utskott, myosinhuvudet, som kan binda till ett intilliggande aktinfilament och bilda en tvärbrygga. Parallellt med filamenten löper längsgående& titinmolekyler, som kopplar ihop Z-linjerna.

Hur dras muskelceller ihop?
När en muskelfiber drar ihop sig, utför myosinhuvudena rörelser, som påminner om årornas rörelser vid rodd. De fäster sig i aktinmolekylerna, bildar en tvärbrygga och viker sig i riktning mot sarkomerens mitt vid M-linjen. Därmed drar de från två håll aktinfilamenten mot M-linjen. Sedan släpper de aktinfilamenten, viker sig åt motsatt håll och upprepar samma rörelser. Det är som när du drar till sig ett rep genom att gripa tag i det med händerna, drar till sig en bit rep, tar ett nytt tag i repet längre fram, drar till sig mer av repet och sedan upprepar rörelserna flera gånger. Notera att aktinfilamenten och myosinfilamenten inte förkortas, när muskelfibern förkortas enligt denna så kallade filamentförskjutningsmekanism.
Tvärbryggornas rörelser gör att sarkomeren förkortas. När många sarkomerer förkortas samtidigt, förkortas muskelfibern. När många muskelfibrer förkortas samtidigt förkortas hela muskeln. Den kontraheras alltså. Tvärbryggornas rörelser drivs av kemisk energi, som utvinns vid nedbrytning av energirika ATP-molekyler till ADP och Pi (fosfat). När ett ATP binds till ett myosinhuvud, bryts ATP ner och överför energi till huvudet. Då laddas huvudet med lägesenergi, viks i riktning mot Z-linjen och stannar där, spänt som en utdragen pilbåge. När huvudet sedan binder till ett aktinfilament, utlöses pilbågen och huvudet viker sig mot M-linjen i sarkomerens mitt.
ATP-molekylerna bildas i muskelfibrerna, när de bryter ner energirika näringsämnen. Vid aerobt arbete bryts glukos och fettsyror från blodet ner med hjälp av syre. Vid anaerobt arbete bryts lagrat glykogen i muskelfibern ner till glukos, som i sin tur bryts ner till mjölksyra utan att syre behövs.
Muskler är elastiska
Sarkomeren innehåller också elastiska element. Den tvärgående M-linjens proteiner håller mysosinfilamenten på plats, men är också elastiska. Titinmolekylerna fungerar som gummiband. De löper från Z-linje till Z-linje och håller därmed ihop sarkomeren. De gör också hela muskeln elastisk. Titin är den största proteinmolekyl vi känner till.
Referenser
- N. Fukuda och S. Ishiwata: Effects of pH on spontaneous tension oscillation in skinned bovine cardiac muscle (Pflügers Archiv 438:125-132, 1999).
- K. E. Barnett, S. M. Barman, S. Boitano, och H .L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- L. Tskhovrebova och J. Trinick: Making muscle elastic, the structural basis of myomesin stretching (PLoS Biol 10: e1001264, 2012).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).

Vid en jämn dragkamp utförs huvudsakligen så kallade isometriska (statiska) muskelkontraktioner. Då trycks musklernas blodkärl ihop så att blodflödet, och därmed syretillförseln, till dem minskar. Muskelcellerna måste då omsätta energi med anaerob, icke syrekrävande cellandning under bildning av mjölksyra. Detta leder till uttröttning, som kan motverkas genom att om möjligt röra på armar och ben.
Muskler är maskiner som kan utföra arbete. Hur regleras den kraft en muskel utvecklar? En muskels längd kan minska, förbli konstant eller öka under en kontraktion. Vad händer vid dessa olika typer av kontraktion? Är muskler elastiska?
En skelettmuskels kontraktion (sammandragning) styrs av muskelns så kallade motoriska enheter. En motorisk enhet består av en motorisk nervcell i det centrala nervsystemet och ett antal muskelfibrer (muskelceller) i musklen. Nervcellen påverkar muskelfibrerna via en nervfiber (axon), som grenar sig inne i muskeln. Grenarna avslutas av så kallade neuromuskulära kontakter med enhetens muskelfibrer. Varje muskelfiber påverkas av en enda neuromuskulär kontakt, som fungerar ungefär som en synaps och verkar på fibern via transmittorn acetylkolin.
Motoriska enheter och muskelkraft
När aktionspotentialer (nervimpulser) skickas längs med den motoriska nervfibern i en motorisk enhet, kontraheras alla dess fibrer. Muskelns kraftutveckling är i varje ögonblick beroende av antalet kontraherande motoriska enheter. Ju fler enheter, och därmed muskelfibrer, som arbetar parallellt med varandra, desto störrre kraft utvecklar muskeln. Om det ingår få muskelfibrer i en muskels motoriska enheter, kan muskelns kontraktioner styras på ett mera finstämt sätt. Musklerna som styr handens rörelser har därför få fibrer i sina enheter, medan ryggmusklerna har många.

Hos en frisk person är musklerna aldrig helt avslappade. Även när du själv slappnar av, så finns det aktiva motoriska enheter i alla skelettmuskler. Alla skelettmuskler utvecklar alltid en mer eller mindre stor kraft, till och med när vi sover eller är medvetslösa.
Alla sarkomerens delar består av proteiner. Den avgränsas på båda sidorna av tvärgående Z-linjer. Dessa linjer ingår också i angränsande sarkomerer, hopkopplade med den ovan avbildade. Flera långsträckta tunna aktinfilament är fästa i Z-linjerna, varje filament bestående av en mängd klotformiga aktinmolekyler. Mellan aktinfilamenten finns tjocka myosinfilament, varje filament bestående av ett stort antal myosinmolekyler. Den tvärgående M-linjen kopplar ihop myosinfilamenten. Varje myosinmolekyl är i sin ena ände försedd med ett utskott, myosinhuvudet, som sticker ut från myosinfilamenet. Huvudet kan binda till ett intilliggande aktinfilament och bilda en tvärbrygga. Parallellt med filamenten löper elastiska längsgående titinmolekyler, som kopplar ihop Z-linjerna.
Ett myosinhuvuds rörelser drivs av energirika ATP-molekyler. När ett ATP binds till ett myosinhuvud, bryts ATP ner och överför energi till huvudet. Då laddas huvudet med lägesenergi, viks i riktning mot Z-linjen och stannar där, spänt som en utdragen pilbåge (Energize i videon). När huvudet sedan binder till ett aktinfilament (Attach) och bildar en tvärbrygga, utlöses pilbågen och huvudet viker sig mot M-linjen i sarkomerens mitt (Tilt).
Isotonisk (koncentrisk) kontraktion
Vid en så kallad isotonisk (koncentrisk) kontraktion utvecklar muskeln kraft, som gör att den förkortas. Dess kraft är således större än den motverkande kraften. Ett exempel är när vi lyfter på en tyngd. Muskelkraften åstadkommes när myosinfilamentens huvuden bildar tvärbryggor, som binder till bindningsställen på aktinfilamenten. Bryggorna viker sig då och utvecklar kraft, drivna av energi från molekylen ATP. De viker sig i riktning mot sarkomerernas mitt, M-linjen. Denna process upprepas flera gånger, ungefär som årtag vid rodd. Aktinfilamenten på båda sidorna av M-linjen förskjutes då längs med myosinfilamenten i riktning mot M-linjen. Detta leder till att sarkomererna, muskelfibrerna och därmed hela muskeln förkortas.

När muskler som på bilden ovan arbetar isotoniskt vid kroppsrörelser, motverkas dock rörelsen i någon mån av (ej utritade) muskler på överarmens baksida, så kallade antagonister, som rätar ut armen. Men de muskler som lyfter hanteln utövar mycket större kraft än antagonisterna. Detta gör att armen kan böjas successivt på ett kontrollerat sätt, ungefär som då du reglerar en bils hastighet med både gas och broms.
Isometrisk (statisk) kontraktion
Vid en så kallad isometrisk (statisk) kontraktion utövar muskeln kraft utan att varken förkortas eller förlängas. Orsken till detta är att muskeln utsätts för en motkraft, som är exakt lika stor som dess egen kraft. Ett exempel är när du försöker lyfta något som du inte kan rubba. Myosinets tvärbryggor utövar då kraft utan att röra på sig, vilket leder till att muskelns längd inte förändras. Myosinbryggorna kan släppa och binda igen upprepade gånger, men de binder varje gång till samma bindningställe på aktinet och flyttar således inte på aktinfilamenten. Men när de är bundna till aktinet utvecklar de kraft.

Vid en isometrisk kontraktion pressas muskelns blodkärl i hop så att dess blodflöde, och därmed dess syretillförsel, minskar eller upphör. Muskeln måste då förlita sig på anaerob, icke syrekrävande cellandning under bildning av mjölksyra. Därför tröttnar den snabbt. Vid en isometrisk kontraktion utvecklar muskeln kraft utan att utföra arbete i fysikalisk mening. Ingen rörelse sker nämligen. Därför omvandlas all energi som muskeln omsätter, till värme, inte alls till mekaniskt arbete.. Detta kan vara svårt att acceptera för den som försöker lyfta en tung sten, men så är det.
Förlängningskontraktion (excentrisk kontraktion)
Vid en så kallad förlängningskontraktion (excentrisk kontraktion) förlängs muskeln, medan den utövar kraft. Orsaken till att den förlängs är att den påverkas av en yttre kraft, som är större än den kraft den själv utvecklar. Den yttre kraften utvecklas vanligen av en annan muskel med motsatt effekt. När vi böjer armen i armbågen utövar de böjande armmusklerna större kraft än de uträtande armmusklerna, men båda muskelgrupperna utövar kraft. Detta är nödvändigt för att vi ska kunna kontrollera rörelsen. Om uträtarmusklerna var helt avslappade, skulle rörelsen bli snabb och fullkomligt okontrollerad. I stället har vi både gas och broms.
Vid en förlängningskontraktion binder myosinets tvärbryggor till bindningsställena på aktinet, utövar kraft och rör sig. Men den yttre kraften är större än den kraft de själva utövar. Därför kan de inte utföra sina ATP-drivna årtag. De tvingas i stället att vika sig i motsatt riktning mot årtagen vid isotonisk kontraktion, bort från M-linjen. De binder upprepade gånger till aktinet och utövar kraft, men de omvända årtagen leder till att aktinfilamenten successivt avlägsnar sig från M-linjen.
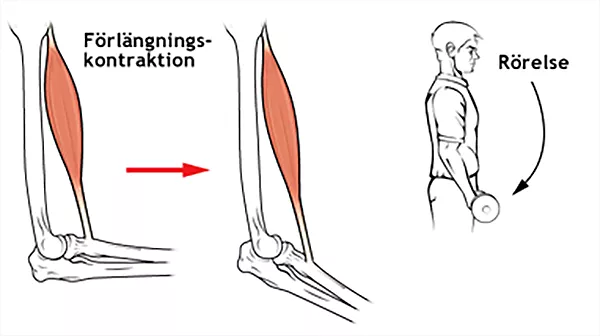
Fördjupning
Ovanstående artikel Muskelkontraktion: hur skelettmuskler drar ihop sig med hjälp av myosin och aktin.
Muskler är elastiska
Tilläggas bör att muskler också har elastiska egenskaper. En fripreparerad muskel, som inte stimuleras av nerver, uppför sig som ett mycket kraftigt gummiband. Den intar ett jämviktsläge. Om du drar ut den och släpper den, så återgår den till jämviktsläget. De elastiska krafterna utövas dels av senor och bindväv, dels av muskelfibrernas sarkomerer genom proteinet titin och M-linjens proteiner. Om en sarkomer sträcks ut så att myosinfilamenten förlorar kontakten med aktinfilamenten, kan den dras ihop igen av dessa proteiner. Elasticiteten ger också ett visst skydd mot överbelastning av musklerna.
Referenser
- K. E. Barnett, S. M. Barman, S. Boitano, och H .L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal physiology (4th ed, Sinauer, 2018 ).
- L. Tskhovrebova och J. Trinick: Making muscle elastic, the structural basis of myomesin stretching (PLoS Biol 10: e1001264, 2012).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (13th ed, McGraw-Hill, 2014).

När mycket hög muskelkraft behövs, använder djur så så kallade pennata, det vill säga fjäderlika, muskler. Sådana muskler finns bland annat i kräftors klor och krokodilers käkar. De kallas så därför att de liknar fåglars fjäderpennor. I pennata muskler löper senan genom hela muskeln och de långsträckta muskelcellerna är fästa i senan som strålarna i en fågelfjäder.
Till vänster på bilden nedan ses några sådana muskler som påverkar fingrarna i människans hand. En fjäderlik muskel utövar stor kraft inom ett litet utrymme, till exempel inuti en kräftklo eller mellan mellanhandsbenen i en människohand. Nedan följer för den intresserade en förklaring till detta fenomen.

Till höger på bilden ovan ses en schematisk bild av en fjäderlik muskel. Senan är svart, muskelfibrerna röda. Muskelfibrerna drar ihop sig i sin längsriktning. Eftersom muskelfibrerna inte löper i muskelns längsriktning kan en fjäderlik muskel inte förkorta sig lika mycket som en vanlig muskel. Dessutom går en del av muskelkraften till spillo. De tjocka blå kraftpilarna svarar mot de krafter fibrerna utvecklar i sin egen längsriktning. Dessa krafter kan delas upp i de krafter som verkar i muskelns längsriktning och förkortar den (tunn blå lodrät pil) och de krafter som drar i senan vinkelrätt mot muskelns längsriktning och därför motverkar varandra (tunna blå vågräta pilar). De vågräta motverkande krafterna är förspilld muskelkraft, som inte leder till någon rörelse. Hittills bara nackdelar således!
Men den totala användbara muskelkraften i en fjäderlik muskel kan trots allt bli större än i en vanlig muskel med samma volym. Muskelkraften är nämligen proportionell mot den totala tvärsnittsytan vinkelrätt mot fiberriktningen. Varför? Jo, ju fler parallella fibrer som drar i samma sena, ju större blir deras totala kraftutveckling. Annorlunda uttryckt, ju fler gubbar som står bredvid varandra och drar i samma rep, ju större kraft kan de utveckla tillsammans.
Till höger på bilden ovan är den totala tvärsnittsytan vinkelrätt mot fiberriktningen i den fjäderlika muskeln proportionell mot den totala längden av alla de fyra gröna sneda dubbelpilarna, medan den i en motsvarande muskel med längsgående fibrer är mycket mindre, proportionell mot längden av den enda bruna vågräta dubbelpilen. En vanlig muskel kan jämföras med ett antal gubbar som står i en korridor och drar i repets ände. Det blir då trångt och väldigt få gubbar får plats. En fjäderlik muskel kan jämföras att gubbarna står längs med hela repet och drar. Då får det plats många fler gubbar i korridoren och tillsammans kan de dra i repet med mycket större kraft. Tack vara den större totala tvärsnittsytan kan således en fjäderlik muskel utöva stor kraft inom ett litet utrymme, men den kan inte förkorta sig lika mycket som en vanlig muskel.

Fördjupning
Vilka djur hoppar högst? Hur kan små djur, som loppor och gräshoppor, hoppa så högt? Se nästa artikel.
Referenser
K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
G. J. Tortora och M. T. Nielsen: Principles of human anatomy (12th ed, Wiley, 2012).

Många djur kan hoppa högt. Här ska jag berätta om några av de bästa höjdhopparna bland landdjuren och, om möjligt, komma fram till vilket djur som hoppar högst. Men först ska jag förklara varför små djur, som loppor och gräshoppor, kan hoppa så högt. Det inofficiella världsrekordet i stående höjdhopp för människor är 190 cm, satt av svensken Rune Almén år 1980, obetydligt högre än cirka 180 cm, vilket är medellängden för män i Sverige. En loppa kan i stående höjdhopp hoppa cirka 80 gånger högre än sin egen kroppslängd på cirka 2-3 mm.
Varför kan insekter hoppa så högt?
Här är några siffror som visar hur högt några hoppande djur kan hoppa. Det handlar om stående höjdhopp, det vill säga hopp utan ansats, så världsrekordet i höjdhopp på 245 cm kommer inte med.
- En loppa hoppar cirka 20 cm
- En knäpparskalbagge hoppar cirka 30 cm
- En gräshoppa hoppar cirka 60 cm
- En människa hoppar cirka 60 cm
Varför hoppar människan bara 60 cm? En specialtränad människa kan hoppa cirka 160 cm i stående höjd, men här handlar det om att förflytta kroppens tyngdpunkt. Vår höjdhoppare har långa ben, så hans tyngdpunkt är cirka 100 cm över marken. Han förflyttar således tyngdpunkten 160 - 100 = 60 cm. Därför hoppar han bara cirka 60 cm. Skillnaderna i hopphöjd mellan de olika djuren är väldigt liten, trots att djuren skiljer sig mycket åt i storlek. Loppan väger cirka 0,5 milligram (mindre än en tusendels gram), skalbaggen cirka 40 milligram (fyra hundradelar av ett gram), gräshoppan cirka 3 gram och människan cirka 70 kg. Människan är 140 miljoner gånger tyngre än loppan och 23 000 gånger tyngre än gräshoppan. Ändå hoppar människan bara ungefär tre gånger högre än loppan och lika högt som gräshoppan. Hur är detta möjligt?

De tre ovan nämnda insekterna är i och för sig speciellt anpassade till att hoppa. Men den lilla skillnaden gentemot människan är ändå märklig. Förenklat kan vi anta, att alla djuren har en likadan kroppsbyggnad. Med hjälp av fysik kan vi då visa följande. Ett litet djur kan utveckla en större kraft per gram kroppsvikt än ett stort. Annorlunda uttryckt så är 23 000 gräshoppor tillsammans mycket starkare än en människa, trots att de tillsammans väger lika mycket som människan. Men en hoppande människa har en mycket längre accelerationssträcka innan avstampet än en hoppande gräshoppa. Därför får människan och gräshoppan ungefär samma utgångshastighet, när de lyfter från marken. Detta gör att de grovt räknat hoppar lika högt. Accelerationssträckan är här den sträcka som den stående hopparens tyngdpunkt rör sig uppåt sig från en hukande ställning tills dess att han lämnar marken. Kom i håg att det handlar om stående höjdhopp utan ansats.
Varför är små djur starkare än stora?
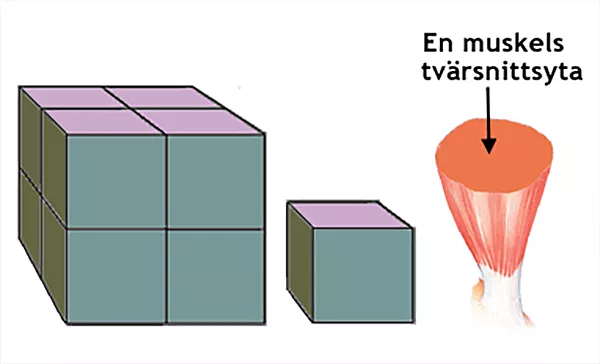
För den intresserade förklaras här utförligt sambandet mellan yta/volym-förhållande och kroppstorlek hos djur. Den lilla kuben i bilden ovan motsvarar ett litet djur, den stora ett stort. Den stora kuben består av åtta små kuber. Det lilla djurets volym, och därmed ungefärligen dess kroppsvikt, är således åtta gånger mindre än det stora djurets. En av den lilla kubens ytor är fyra gånger mindre än en av den storas ytor. Alltså är den lilla kubens totala yta fyra gånger mindre än den storas. Det lilla djurets yta är således bara fyra gånger mindre än det storas. Detta gäller inte bara kroppsytan, utan också alla andra ytor i kroppen, som musklernas tvärsnittsytor. Om en muskels tvärsnittsyta är större kommer fler muskelceller att arbeta parallellt med varandra och dra i dess sena. Därmed utvecklar muskeln en större kraft. Ju fler gubbar (muskelcellerna) som drar i samma rep (senan), ju större kraft utvecklar de. Det lilla djuret väger således åtta gånger mindre än det stora, men har bara fyra gånger mindre tvärsnittsytor i sina muskler. Små djur är således starkare än stora i förhållande till sin kroppsvikt. Detta är förklaringen till att myror och andra insekter lätt kan bära bördor, som väger mer än de själva. Det ger också små djur en stor fördel när de hoppar. Resonemanget ovan är en tillämpning av en geometrisk sats formulerad redan av den gamle greken Euklides: volymskalan är lika med längdskalan i kubik och ytskalan är längdskalan i kvadrat.
Hoppande insekter fungerar som armborst
Höga hopp och övriga mycket snabba rörelser hos insekter och andra leddjur utförs vanligen inte direkt genom att muskler drar ihop sig. Muskelkontraktioner är faktiskt för långsamma. I stället spänns elastiska strukturer som fungerar som gummiband. Dessa strukturer låses sedan i spänt läge. Rörelsen utlöses av en avtryckarmekanism, som leder till att de elastiska strukturerna mycket snabbt förkortas och påverkar ben eller andra kroppsdelar. Insekterna fungerar alltså som slangbågar eller, med en bättre liknelse, armborst. De elastiska strukturerna kan vara delar av det yttre skelettet (kutikulan), senliknande bildningar eller musklerna själva. Ofta innehåller de det mycket elastiska proteinet resilin, som har bättre elastiska egenskaper än gummi.

Till dessa eminenta hoppare kan vi lägga den cirka 6 mm långa spottstriten Philaenus spumarius, som kan utföra höjdhopp på 70 cm eller 115 kroppslängder med hjälp av bakbenen. Den accelererar så snabbt att den utsätts för en G-kraft, som är mer än 500 gånger större än tyngdkraften. En erfaren pilot utan skyddande så kallad G-dräkt kan klara 7 G, men bara under en mycket kort period. Sedan drabbas hen av medvetslöshet.

Hoppande däggdjur
Människan är inte speciellt väl anpassad till höjdhopp. Det finns många däggdjur som är anpassade till att hoppa och som kan hoppa betydligt högre än människan både i stående höjdhopp och i hopp med ansats. Ett sådant djur är dvärggalagon (Galago senegalensis) som kan hoppa mycket högt i stående höjdhopp. Galagoerna är halvapor. Det finns flera galagoarter som lever i träden i den afrikanska skogen. En dvärggalago som väger cirka 250 gram kan hoppa cirka 225 cm i stående höjdhopp. Den slår både loppan och människan med bred marginal.
En sådan galago skulle ha platsat i OS i höjdhopp och detta utan att utnyttja ansatsbanan. Observera dock att både människor och galagoer hoppar med båda benen vid stående höjdhopp, medan en olympisk höjdhoppare som använder ansats bara får hoppa med ett ben. Med nuvarande höjdhoppsregler hade galagon således blivit diskad på OS. Å andra sidan gynnas de mänskliga höjdhopparna på ett otillbörligt sätt genom att deras tyngdpunkt ligger åtminstone en meter över galagons. Ofta lyfter de dessutom inte sin tyngdpunkt över ribban, även om de klarar höjden. De böjer ju kroppen, så att överkroppen och underbenen ligger under ribban, då de når hoppets högsta punkt. Detta kan leda till att kroppens tyngdpunkt ligger nedanför kroppen och passerar under ribban.
Varför är dvärggalagoerna så bra på att hoppa? Den viktigaste orsaken är att de har hoppmuskler som upptar cirka 10 procent av kroppsvikten. Viktigt är också att de har långa bakben, som genom en extra stor hävstångseffekt kan ge dem hög utgångshastighet vid hoppet. För den intresserade förklaras hävstångseffekten i faktarutan nedan. Även de andra medlemmarna av familjen Galagidae är utomordentliga höjdhoppare. De är också bra längdhoppare och påstås kunna prestera hopplängder på upp emot 12 meter. Galagoerna finns i Afrika.

Sådana hävstångsmekanismer i skelettet utnyttjas av djur, som springer snabbt och hoppar högt. Det är därför som sådana djur har långa ben. Fördelen med hävstängerna är att de ger en snabb rörelse med stort utslag. Nackdelen med dem är att kraften blir mindre. Detta kompenseras av dessa djur genom att de har extra starka muskler med stor tvärsnittsyta (se bilden högre upp på sidan). Det senare gäller i synnerhet hoppande djur, som inte bara måste ta sig framåt, utan också uppåt och övervinna tyngdkraften.
För den matematiskt intresserade kan vi beskriva förloppet med hävstångslagen: LA x FA = LB x FB. LA (cm) är delen A:s längd, LB (cm) delen B:s. FA (N) är den kraft muskeln utvecklar på A, FB (N) den kraft B:s ände utvecklar mot underlaget.
De sydostasiatiska spökdjuren (släktet Tarsius) är också mycket bra hoppare. Spökdjuren har extremt långa ben och hoppmuskler som upptar en mycket stor andel av kroppsvikten. De utgör en särskild grupp bland primaterna, jämte halvapor och apor. En spökdjursart uppges göra mer än 5 meter långa och upptill 1,5 meter höga hopp, trots att den bara väger 80–160 gram.
Precis som insekter, utnyttjar däggdjur elastiska delar i rörelseapparaten, när de hoppar och även när de springer. Vid fotens nedslag på underlaget sträcks dessa elastiska delar som gummiband och lagrar elastisk lägesenergi. Vid avstampet omvandlas lägesenrgin till rörelseenergi som, jämte muskelsammandragningar, bidrar till att lyfta djuret från underlaget och föra det framåt. De elastiska delarna kan vara senor, bindvävsskikt och till och med muskler. Löparskor med elastiska sulor och löparbanors underlag har liknande effekter. Rörelser med avtryckarmekanismer, som hos insekter, finns dock inte hos däggdjur.

Det finns fler däggdjur som kan hoppa högt, men det är svårt att hitta säkra uppgifter. Det finns uppgifter om att vissa små antiloper kan göra höjdhopp på 2–3 meter, oklart om det är med eller utan ansats. Notera dock att de, med sina långa ben, har en tyngdpunkt som ligger högt över marken. Jättekängurur uppges enligt osäkra uppgifter kunna göra höjdhopp på 1,8–3 meter (olika källor), förmodligen med ansats. Fältharen (Lepus europaeus) uppges enligt en äldre osäker uppgift kunna göra höjdhopp på 4,5 meter och den vitsvansade åsneharen (Lepus townsendii) höjdhopp på otroliga 6,4 m, även här osäkert om det är med eller utan ansats. Enligt en annan osäker uppgift kan pumor (Puma concolor) göra höjdhopp på 5,5 m. Det måste betraktas som oklart vilket djur som innehar höjdhoppsrekordet bland däggdjuren. Vi får dock intrycket att det är få, om några, däggdjur som kan hoppa högre än 3 m.
Fördjupning
Varför är kräftans klor och krokodilens käkar så starka? Om fjäderlika muskler. Artikeln finns precis ovanför.
Varför kan vi svimma när vi står upp? Hur påverkar tyngdkraft och g-krafter ormar och astronauter? Du hittar artikeln under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
Referenser
- Anonymous: Red Kangaroo (Zoos Victoria, hämtad 19 september 2016).
- Anonymous: Red Kangaroo, Macropus rufus (National Geographic Society, hämtad 19 september 2016).
- Anonymous: Western Tarsier (Tarsius bancanus) (Sarawak National Park, hämtad 19 september 2016).
- J. Brackenbury och H. Hunt: Jumping in springtails: mechanism and dynamics (Journal of Zoology 229:217-236, 1993).
- M. Burrows: Jumping performance of froghopper insects (Journal of Experimental Biology 209:4607-4621, 2006).
- M.-C. Cadiergues et al.: A comparison of jump performances of the dog flea, ;Ctenocephalides canis (Curtis, 1826) and the cat flea, Ctenocephalides felis felis (Bouché, 1835) (Veterinary Parasitology 92:239–241, 2000).
- R. H. Crompton och W. I. Sellers: A consideration of leaping locomotion as a means of predator avoidance in prosimian primates (In: Primate anti-predator strategies, (K. A. I. Nekaris and S. L. Gursky, ed., pp. 127-145, 2007, Springer, New York).
- P. Crona: Rekorden som inte är (Göteborgs Friidrottsförbund, hämtad de 19 september 2016).
- E. Davies: The greatest jumper on Earth is probably not a flea (BBC Earth, uppdaterad 26 augusti 2016).
- T. Dewey: Macropus rufus, red kangaroo (Animal Diversity Web, hämtad 15 september 2016).
- E. C. B. Hall-Craggs: An analysis of the jump of the lesser galago (Galago senegalensis) (Journal of Zoology 147:20-29, 1965).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. Schmidt-Nielsen: Animal physiology (5th ed., Cambridge University Press, 1997).
- S. Sudo et al.: Observations on the springtail leaping organ and jumping mechanism worked by a spring (Journal of Aero Aqua Bio-mechanisms 3:92-96, 2013).
- S. Sudo et al.: The kinematics of jumping of globular springtail (Journal of Aero Aqua Bio-mechanisms 3:85-91, 2013).
- G. P. Sutton och M. Burrows: Biomechanics of jumping in the flea (Journal of Experimental Biology 214:836-847, 2011).
- B. Wyrick och J. R. Brown: Acceleration in aviation: G-force (Federal Aviation Administration, hämtad 19 september 2016).

Vi har väl alla påträffat insekter, särskilt skalbaggar, som ligger på rygg och kämpar länge för att vända sig rätt. Klarar de av detta och, i så fall, hur gör de? Det naturliga urvalet borde, tycker jag, obönhörligt sortera bort de insekter som inte kan vända sig på rätt köl.
Insekter som hamnar uppochned är inte dömda att dö. Alla insekter, även skalbaggar, har upprättningsreflexer med hjälp av vilka de kan komma på rätt köl när de hamnar på rygg. De klarar av att rätta upp sig även på plana underlag, med hjälp av benen eller av vingarna. Skulle de i enstaka fall misslyckas, beror det förmodligen mestadels på att de har nedsatt livskraft. De svåraste underlagen, glatta vågräta ytor, är dessutom mycket sällsynta i naturen. Studiet av insekters upprättningsreflexer är av stort tekniskt intresse och kan leda till bättre vandrande robotar. Insektsliknande sexbenta robotar klarar sig mycket bättre än tvåbenta i obanad terräng. Men vi vill ju inte att de ska vara hjälplösa, om de hamnar på rygg.
Hur vänder sig insekter rätt från ryggläge?
När en skalbagge ligger på rygg, ser det ut som om den rör benen på måfå. Detta är långt ifrån fallet. Det handlar i stället om komplicerade nervösa reflexer. Olika arter kan använda olika reflexer, beroende på benens längd och kroppens form. Till en början letar baggen vanligen efter ett fäste i underlaget för något eller några av benen. Finner den ett fäste kan den rätta upp sig på flera olika sätt med hjälp av benmuskulaturen. Den kan vrida kroppen sidledes i riktning mot det förankrade benet. När kroppens tyngdpunkt passerat över dess anläggningspunkt på underlaget, ramlar baggen automatiskt ner på rätt köl. Den kan också lyfta kroppen med hjälp av ett ben på ena sidan, så att kroppen rullar över sidledes åt motsatt håll.
Vissa skalbaggar med långa ben kan lyfta upp kroppen med hjälp av ett ben eller ett benpar, så att bakänden vilar på underlaget och sedan göra en kullerbytta. När kroppen tyngdpunkt passerar över bakänden, ramlar baggen framåt och kan sedan traska vidare. Andra skalbaggar böjer kroppen i en sprättbåge så att bara huvudet, bakänden och ett ben vilar på underlaget. Sedan ramlar de ner på sidan och kan då lätt rätta upp sig. Det finns fler upprättningsmetoder, både hos skalbaggar och hos andra insekter.

Gregor Samsa drabbas dock av ångest och plågor efter sin metamorfos. Familjen vänder sig mot honom och han känner sig värdelös. Novellen kan, liksom andra mästerliga litterära verk, tolkas på många olika sätt. Den kan handla om tillvarons absurditet, människors förakt för andras bräcklighet, individens främlingskap i samhället, vår oförmåga att leva upp till andras förväntningar, allt detta på en gång eller något helt annat.
Den märkligaste upprättningsreflexen hittar vi hos knäpparskalbaggarna. De kan utan hjälp av benen hoppa mycket långt upp i luften. Sedan har de 50 procents chans att hamna på rätt köl. Om de hamnar fel, gör de om manövern.
Även om det tar lite tid för en del arter, kan skalbaggar liksom andra insekter således vända sig rätt. Annars hade skalbaggarna inte blivit jordens framgångsrikaste djurgrupp, om vi ser till antalet kända arter.
Referenser
- M. Davis et al.: A review of self-righting techniques for terrestrial animals (International Workshop on Bio-inspired Robots, École des Mines de Nantes, France), April 6th-8th, 2011).
- L. Frantsevich: Righting kinematics in beetles (Insecta: Coleoptera) (Arthropod Structure & Development 33:221-235, 2004).
- F. Kafka: Förvandlingen (Bakhåll förlag, 2008; översatt av Hans Blomqvist och Erik Ågren).
- S. Sommelius: Otaliga tolkningar av ohyra (Helsingborgs Dagblad, 10 januari 2005).

Fotografiet visar en pärlbåt (släktet Nautilus), den sista kvarlevande av en fordom blomstrande stam. Nautilus är en bläckfisk med yttre skal. Skalet har en intressant funktion. Det är fyllt med luft och ger Nautilus hjälp att hålla sig svävande i vattnet.
Nautilusarterna lever i Stilla havet och Indiska oceanen. Flera grupper av bläckfiskar med raka eller spiralformade skal var vanliga under tidigare geologiska epoker och deras skal återfinns ofta som fossil ("ortoceratiter", ammoniter och belemniter). Nautilus är den enda nu levande bläckfisken med yttre skal. Hos andra nutida bläckfiskar med skal finns skalet inne i djuret och syns inte utifrån.

Bläckfiskarnas skal fungerar (precis som fiskarnas simblåsa) som en flythjälp. Skalet innehåller nämligen kammare som bläckfisken fyller med luft. En kammare är först fylld med vävnadsvätska. Bläckfisken pumpar sedan med en energikrävande process ut löst salt, natriumjoner och kloridjoner, ut ur kammaren. Den koncentrationsskillnad, som då uppkommer, gör att vatten följer efter saltet genom så kallad osmos. När vattnet försvinner, sjunker trycket i kammaren, vilket leder till att bubblor av syrgas och kvävgas bildas, precis som trycket sjunker och bubblor av koldioxid bildas, när du öppnar en flaska med läsk. Till slut är nästan hela kammaren fylld med luft.
Fördelen med att ha ett skal som flythjälp är att det inte, som en simblåsa, pressas ihop av trycket (som ju ökar med djupet under havsytan). En nackdel med skal är att de väger mycket mer än simblåsor. En annan nackdel med skal att de inte fungerar på stora djup, där vattentrycket blir så stort att skalet krossas. Fiskar med simblåsor kan fylla simblåsorna med gas upp till flera hundra atmosfärers tryck, så att de motstår vattentrycket på mycket stora djup. Men även för fiskarna finns det en nedre gräns, under vilken simblåsor inte fungerar.
Referenser
Texten har uppdaterats och utökats år 2013.
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).
- P.C. Withers: Comparative Animal Physiology (Saunders College Publishing, 1992).
Kan alla däggdjur simma? Jag kan naturligtvis inte ge ett säkert svar på denna fråga. Simkunnigheten har bara testats hos ett fåtal av alla landlevande däggdjursarter. Men vi kan ändå diskutera frågan utifrån den kunskap som finns.
En del tror att det finns däggdjur som inte kan simma. Andra menar att alla däggdjur kan simma. Akvatiska djur, till exempel sälar och valar, simmar och dyker naturligtvis bra, liksom halvakvatiska djur, till exempel bävrar, bisamråttor och uttrar.
De flesta däggdjur kan troligen simma spontant
Problemet gäller landdäggdjuren. Många av dem simmar spontant, utan att behöva lära sig. I vattnet använder de i regel samma gångarter som de använder på land med benen rakt under kroppen, så kallat hundsim. Så vitt jag kunnat se har vi, med fåtal undantag, inte hittat några landdäggdjur som saknar förmågan att simma spontant. Men en mängd djur har ju inte testats. Undantagen är giraffen, människan och människoaporna. Giraffens simkunnighet är oklar. Människor måste lära sig att simma. Spädbarn kan hålla andan under vattnet ("babysim"), men de simmar inte upp till ytan. Forskare har nu också visat att schimpanser och orangutanger kan lära sig att simma. Intressant nog simmar de inte hundsim. Schimpansers armrörelser påminner om crawl, deras benrörelser om bröstsim.
Varför måste människor lära sig simma?
Varför måste människor lära sig att simma? En möjlig orsak är vår upprätta kroppsställning. När fyrfota däggdjur ligger i vattnet, hamnar kroppen i vågrätt läge. Då blir det lättare för dem att simma med hjälp samma gångarter som används på land. En människa som hamnar i vattnet har svårare att hålla huvudet över vattenytan och måste använda andra rörelsemönster vid simning än vid gång på land.
Tre osannolika simmare
Förvånande nog kan fladdermöss simma genom att slå med vingarna. Och elefanter, är utmärkta simmare, som använder snabeln som snorkel. Till och med sengångare kan simma.
Om vattenapeteorin (littoralhypotesen)
Enligt den så kallade vattenapeteorin eller littoralhypotesen har människan under sin evolution levt nära vatten, anpassat sig till att simma och dyka och därigenom kunnat utnyttja nya näringskällor. Det finns anatomiska och fysiologiska argument för denna teori, men också många sådana argument som talar emot den. Teorin har inte vunnit genklang. Fossilfynden tyder på att de olika människoarterna under loppet av sin evolution levt i ett varierande landskap med varierande klimat, tidvis i skog, tidvis på savann och tidvis i ett blandat landskap. Men det finns också fossilfynd som visar att människoarter, åtminstone vissa populationer under vissa perioder, levde i närheten av vatten och hämtade föda därifrån. Med tanke på den begränsade kunskap som finns om människans evolution kan vi inte utan vidare förkasta vattenapeteorin.
Referenser
- R. Bender och N. Bender: Swimming and diving behavior in apes (Pan troglodytes and Pongo pygmaeus). First documented report (American Journal of Physical Anthropology 152:156-162, 2013).
- A. I. Dagg och D. E. Windsor: Swimming in northern terrestrial mammals (Canadian Journal of Zoology 50:117-130, 1972).
- J. Gabbatiss: The strange experiments that revealed most mammals can swim (BBC Earth, 21 mars 2017).
Plattmaskar
Videon ovan visar hur virvelmaskar (planarier) rör sig. Dessa plattmaskar glider fram över underlaget på en yta av slem som de själva producerat. Den framåtdrivande kraften åstadkoms av en mängd cilier (flimmerhår) som sitter på buksidans hudceller. Virvelmaskar har också under huden ett muskellager med cirkulärt anordnade fibrer och ett med längsgående fibrer. Muskulaturen kan, i synnerhet hos stora virvelmasksarter, hjälpa till att driva masken framåt. Innehållet i masken fungerar då som ett så kallat hydrostatiskt skelett, en vätskebehållare som kan ändra form, men inte volym. En mask kan göra kroppen längre och smalare, när den drar ihop den cirkulära muskulaturen, eller kortare och tjockare, när den drar ihop den längsgående muskulaturen. Tendenser till ett sådant rörelsemönster syns i filmen.
Daggmaskar
Videon ovan visar hur en daggmask förflyttar sig. Daggmaskar har ett hydrostatiskt skelett och förflyttar sig enbart med hjälp av cirkulär och längsgående muskulatur, inte med cilier. När maskens bakände har fäste i underlaget och framänden förlängs och smalnar, så kommer framänden att transporteras framåt. Om framänden sedan får fäste i underlaget och bakänden förkortas och tjocknar, så kommer bakänden att transporteras framåt. Hos daggmaskar underlättas rörelserna av att maskarna är segmenterade och försedda med borst. Segmenteringen gör att kroppen blir uppdelad i flera vätskebehållare som ligger i rad och kan arbeta oberoende av varandra. Borsten ger masken fäste mot underlaget.
Det "kameraskygga" djuret, som hoppar ut ur bilden när pratbubblan dyker upp, är en hoppstjärt (ordningen Collembola bland insekterna). Hoppstjärtarna är en ursprunglig insektsgrupp med egenartad uppbyggnad. De har inte vingar och är försedda med en hoppgaffel på undersidan av bakkroppen. När de sprätter ut hoppgaffeln kan de hoppa upptill 10 cm. I den övre delen av marken, förnaskiktet, kan det kan förekomma mer än 1 000 exemplar i en liter förna. De lever bland annat på multnande växtdelar och svampmyceler.
Referenser
R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2nd ed, 2003).

Hur gick det till när fåglarna började flyga? Den evolutionära utvecklingen från en reptil, som går på fyra ben, till en fågel som behärskar luftrummet, förefaller mycket drastisk. Men utvecklingen skedde stegvis under lång tid. En del dinosaurier utvecklade fjädrar, men inte till att flyga med. Sedan utvecklades frambenen, kanske också bakbenen, till vingar med flygdugliga fjädrar. Därefter började fåglarnas förfader segelflyga. Till slut omvandlades rörelseapparaten och fjäderdräkten så att aktiv flygning med två vingar blev möjlig. Fåglarna är ensamma bland ryggradsdjuren om att kunna röra sig såväl på marken som i luften och i vattnet, låt vara att alla fåglar inte behärskar simkonsten.
Aktiv flygning har bara utvecklats hos fyra djurgrupper. Insekterna var först. De tidigaste flygande insekterna uppträdde under karbonperioden (cirka 360–300 miljoner år före nutid). Pterosaurierna ("flygödlorna") uppträdde först under triasperioden (cirka 250–200 miljoner år före nutid) och dog ut under kritaperioden (cirka 145–70 miljoner år före nutid). De tidigaste fåglarna uppträdde under juraperioden (cirka 200–145 miljoner år före nutid), förfadern till alla nutida fåglar dock betydligt senare. Fladdermössen uppträdde troligen i början av paleogenperioden (cirka 66–23 miljoner år före nutid).
Fåglarnas ursprung
Notera att fåglarna numera betraktas som en undergrupp bland dinosaurierna. Fåglarna var de enda dinosaurier som överlevde den stora katastrofen för cirka 70 miljoner år sedan, då många djurgrupper dog ut. Orsaken till utdöendet var en jättestor meteorit, som träffade jorden, enorma vulkanutbrott i Indien eller båda dessa händelser tillsammans. Efter utdöendet gick fåglarna en lysande framtid till mötes och utvecklades till alla de fågelgrupper som lever i dag.
Fåglarna utvecklades ur en grupp små dinosaurier som gick på två ben. De tillhörde gruppen Theropoda tillsammans med bland annat Tyrannosaurus rex. Dessa dinosaurier var försedda med fjädrar. De tidigaste fjädrarna var dunlika och gick inte att flyga med. De fungerade kanske i stället som värmeisolering. Dessa dinosaurier kan nämligen ha varit jämnvarma. I så fall höll de en konstant kroppstemperatur högre än omgivningens, precis som dagens däggdjur och fåglar. En annan möjlighet är att en färgad fjäderdräkt möjliggjorde signalering mellan individer inom arten, precis som hos många av dagens fåglar.

Hos vissa dinosaurier utvecklades fjädrar som påminner om nutida fåglars vingpennor på frambenen, hos andra på både frambenen och bakbenen. Sådana fjädrar kunde fungera som bärplan vid flygning. Detta är ett exempel på så kallad exaption. Exaption innebär att strukturer eller organ, som utvecklats med en funktion, i det här fallet gångben och fjädrar, får en helt annan funktion. Exaption är en mycket vanlig evolutionär mekanism.
Hur utvecklade fåglarna sin flygförmåga?
Det finns flera teorier om hur fåglarna började flyga. Alla teorier utgår från att glidflygning föregick aktiv flygning. Följande två teorier är de populäraste:
Enligt "från träden och nedåt"-teorin levde de i träd och hoppade från gren till gren. Så småningom började de segelflyga mellan träden och fick vingar. Ännu senare började de flaxa med vingarna och utvecklade aktiv flygning.
Enligt "från marken och uppåt"-teorin sprang dinosaurierna på marken och hjälpte till med vingarna, när de gjorde långa hopp eller när de sprang uppför sluttningar. Så småningom började de segelflyga. Ännu senare började de flaxa med vingarna och utvecklade aktiv flygning.
Det är emellertid svårt att bekräfta hypoteserna. Studier av de aerodynamiska egenskaperna hos modeller av fossila dinosaurier pågår och kan leda till intressanta resultat. Detsamma gäller studier av nutida fåglars utveckling som foster och ungar. Det står emellertid klart att flygförmågan utvecklats gradvis, med flera funktionella mellansteg. I dag finns det många trädlevande djur inom flera olika djurgrupper som utvecklat segelflykt, men inga markbundna. Detta ger visst stöd åt "från träden och nedåt"-teorin.
Fjädrarnas uppbyggnad
Som en anpassning till flygning utvecklades de dunlika fjädrarna till sådana fjädrar som dagens fåglar har på vingarna och stjärten, vingpennor och stjärtpennor, se bilden ovan. På flygfjädrar utgår strålar från skaftet och från strålarna utgår bistrålar. Hakar på bistrålarna håller ihop strålarna så att fjädern bildar ett så kallat fan, en skiva som kan utöva kraft gentemot luften. Vingfjädrar som används till att flyga med måste vara asymmetriska, det vill säga fanet måste vara smalare på ena sidan om skaftet än på den andra. Framkanten på en vingfjäder måste nämligen vara smalare än bakkanten, annars är den inte stabil i en framifrån kommande luftström. En viktig anpassning till flygning hos dagens fåglar är att svansen förkortats till en så kallad pygostyl och ersatts av stjärtpennor. Stjärtpennorna gör det lättare för fåglarna att manövrera i luften. Dagens fåglar har fortfarande närmast huden kvar dunfjädrar som framför allt bidrar till deras värmeisolering.
Forskarna trodde länge att fjädrar utvecklades direkt ur de fjäll och plåtar som finns hos många reptiler och även på benen hos dagens fåglar. Numera menar de flesta dock att fjädrarna är en nybildning som uppträdde hos dinosaurierna och således inte omvandlade fjäll eller plåtar. Dock finns det hos ryggradsdjuren grundläggande genetisk mekanism i huden genom vilka olika specialiserade hudstrukturer kan utvecklas ur förtjockade överhudsstrukturer, så kallade plakoder. Troligast är att kräldjursfjäll, fågelfjädrar och däggdjurshår utvecklades parallellt med varandra, men att de alla härstammar från en hudstruktur som fanns hos en gemensam förfader till kräldjuren, inklusive fåglarna, och däggdjuren. Denna hudstruktur bildades under fosterutvecklingen ur en speciell typ av plakod, som senare under evolutionen utvecklades i olika riktningar.

Hur skelett och muskler anpassats till flygning
En viktig struktur som finns hos alla nutida flygande fåglar är den kraftiga bröstbenskammen (carina). Den är nödvändig för skickliga flygare, eftersom den ger ett utökat fäste för de kraftiga muskler (m. pectoralis) som sänker vingarna och ger fågeln lyftkraft. En stor bröstbenskam gör att dessa flygmuskler kan vara tjockare och därmed utöva större kraft. Bröstbenskammen fungerar faktiskt också som fäste för de muskler som höjer vingarna (m. supracoracoideus). Dessa musklers senor löper i en båge över korpbenet i skuldergördeln och fäster i överarmsbenets översida. De drar därför vingarna uppåt. Samma mekanism använder du, då du lyfter en tyngd genom att dra nedåt i ett rep, som är fäst i tyngden och som löper över en stång ovanför den. "Urfågeln" (Archaeopteryx, se bilden överst på denna sida) saknade bröstbenskam och dess korpben hade en annan utformning än hos nutida fåglar. Det är därför oklart om Archaeopteryx kunde flyga aktivt.
Skelettbenen hos nutida fåglar är till största delen fyllda med luft, inte med benmärg som hos däggdjuren. Detta gör kroppen lättare, något som underlättar flygning. Även Archaeopteryx hade luftfyllda skelettben. Tvärgående balkar inuti hålrummet ökar benens hållfasthet. Utskott från de så kallade luftsäckarna finns inuti benen hos dagens fåglar. Luftsäckarna ingår i fåglarnas avancerade andningsapparat.
Är "urfågeln" fåglarnas urfader?
Archaeopteryx har länge placerats vid basen av fåglarnas stamträd. Forskare har emellertid numera hittat en rad, mer eller mindre fågellika, fossila dinosaurier med fjädrar, särskilt i Kina. Många är yngre än Archaeopteryx, några är äldre. Fjädrarna har olika utseende. En del av dessa djur har fjäderpennor på frambenen, andra på både frambenen och bakbenen. En del menar till och med att fåglarnas förfader flög med både frambenen och bakbenen och att bakbenens fjäderpennor senare tillbakabildades. Nya studier av fossilen tyder faktiskt på att Archaeopteryx hade fjäderpennor på benen och kanske var fyrvingad. Archaeopteryx läge på fåglarnas stamträd måste numera betraktas som mera oklart. Den står kanske inte så nära de nutida fåglarnas urfader, som forskarna tidigare trott. Den skulle kunna representera en sidogren bland dinosaurierna, som inte ledde direkt fram till dagens fåglar.
Fåglar som förlorat flygförmågan
Det finns i dag ett flertal fåglar som har förlorat flygförmågan helt eller flyger mycket dåligt och sällan. De har kvar vingarna, men dessa är mer eller mindre förkrympta. Hit hör de strutsartade fåglarna: strutsen i Afrika, nanduerna i Sydamerika, emun i Australien, kasuarerna i Australien och på Nya Guinea, kivierna på Nya Zeeland samt de relativt nyligen utrotade moafåglarna på Nya Zeeland och elefantfåglarna på Madagaskar. Med få undantag, till exempel kivierna, är eller var de förmodligen för stora för att kunna flyga och dessutom väl anpassade till att röra sig på marken.
Hos pingvinerna har vingarna omvandlats till fenliknande simredskap. De använder samma nedärvda rörelsemönster som flygande fåglar, när de simmar. De "flyger" alltså genom vattnet. Detta är ytterligare ett utmärkt exempel på så kallad exaption.

Många fåglar som lever på öar kan inte flyga. Det anses bero på att det saknats predatorer i deras miljö och kanske också på att flygande landfåglar riskerar att blåsa bort från sin ö vid kraftiga stormar. Flera av dessa fåglar har utrotats i sen tid, ofta på grund av att predatorer förts till öarna av människan. Ofta har det varit råttor som ätit upp fåglarnas ägg. Så var förmodligen fallet med den kanske mest berömda av dessa fåglar, dronten på ön Mauritius öster om Madagaskar, se bilden ovan. En lika stor släkting till dronten, solitären, utrotades på den närbelägna ön Rodrigues.
Påfallande många ölevande arter inom familjen rallar har förlorat flygförmågan. Många av dessa fåglar har utrotats, men många finns också kvar, ofta på avlägsna små öar i världshaven. De är nästa alltid endemiska för sin ö, vilket innebär att de bara finns (eller har funnits) där. Ett exempel är den utrotningshotade takahen på Nya Zeeland. Något bättre har det gått för wekarallen, även den på Nya Zeeland. Den kan försvara sig mot råttor och till och med äta upp dem. Rallfamiljen representeras i Sverige bland annat av sothönan och rörhönan.
Fördjupning
Hur stor kan du vara och ändå flyga? Om stora fåglar och flygödlor. Du hittar artikeln under rubriken Varför är vissa djur stora medan andra är små?.
Fåglarnas färgrika fjädrar och däggdjurens hårstrån: hur de bildas och får sin färg. Pigment och strukturfärg. Artikeln finns under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?.
Referenser
- J. M. Clark: Dinosaurs and The origin of birds (In: eLS, John Wiley & Sons, Chichester, 2013).
- R. A. Close och E. J. Rayfield: Functional morphometric analysis of the furcula in mesozoic birds (PLoS ONE 7: e36664).
- D. Dhouailly et al.: Getting to the root of scales, feather and hair: As deep as odontodes? (Experimental Dermatology 1-6, 2017).
- N. Di-Poï och M. C. Milinkovitch: The anatomical placode in reptile scale morphogenesis indicates shared ancestry among skin appendages in amniotes (Science Advances 2:e1600708, 2016).
- G. Dyke et al.: Aerodynamic performance of the feathered dinosaur Microraptor and the evolution of feathered flight (Nature Communications 4:2489, 2013).
- P. Godefroit et al.: A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds (Nature 498:359-362, 2013).
- M.J. Greenwold och R.H. Sawyer: Linking the molecular evolution of avian beta (β) keratins to the evolution of feathers (Journal of Experimental Zoology B 316:609–616, 2011).
- G. E. Goslow, Jr: The avian shoulder: an experimental approach (American Zoologist 29:287-301, 1989).
- A. M. Heers: Evolution of Avian Flight (In: eLS, John Wiley & Sons, Chichester, 2013).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- X. Xu: Evolution: Taking wing with weak feathers (Current Biology 22: R992-R994, 2012).
- X. Zheng et al.: Hind wings in basal birds and the evolution of leg feathers (Science 339:1309-1312, 2013).
Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten

Däggdjur och fåglar har som bekant hjärtan med fyra rum, två förmak och två kammare. Mindre bekant är att krokodilerna också har ett fyrarummigt hjärta med två förmak och två kammare. Alla andra nu levande reptiler och alla groddjur har ett trerummigt hjärta med två förmak och en kammare. Det är en stor skillnad mellan att ha tre rum och att ha fyra rum, men inte så som det ofta sägs i skolböckerna. Och det kan till och med vara fördelaktigt att ha ett trerummigt hjärta.
Fyrrummiga hjärtan hos däggdjur och fåglar
Hos däggdjur och fåglar finns två separata kretslopp som är kopplade i serie efter varandra. Se på bilden nedan. Syrgasrikt blod pumpas ut ur vänster kammare via aortan till kroppens organ. Detta blod, som nu är syrgasfattigt, kommer tillbaka från organen till höger förmak. Det pumpas sedan av höger kammare via lungartären till lungorna där det tar upp syrgas. Blodet, som nu är syrgasrikt, flyter därefter från lungorna till vänster förmak och sedan till vänster kammare. Nu är cirkeln sluten.

Vårt hjärta är alltså inte en pump, utan två pumpar Den vänstra och den högra hjärthalvan är separata pumpar. De båda pumparna arbetar synkront, det vill säga de slår samtidigt. De pumpar också ut samma volym blod per tidsenhet. Det senare är en nödvändig konsekvens av att de båda pumparna och de båda kretsloppen alla är kopplade i serie med varandra. Se på bilden ovan. Om höger kammare pumpade ut mer blod per tidsenhet än vänster skulle ju blodvolymen i lungkretsloppet hela tiden öka och volymen i kroppskretsloppet hela tiden minska! Och vice versa. Hos en människa i vila pumpar de båda kamrarna ut vardera cirka 5 liter blod per minut. Hos hårt arbetande elitidrottsmän kan flödet ur varje kammare öka till 35 liter per minut eller mer. Detta är ett betydligt större flöde än det du får om du skulle vrida på kökskranen därhemma på max! Men gör inte det, du kommer att stänka vatten över hela köket.
Trerummiga hjärtan hos groddjur och reptiler
Hos grodor och de flesta reptiler pumpar samma kammare blod till både lungorna och till kroppens organ. Se på bilden ovan. Blodet från båda kretsloppen kommer sedan tillbaka till denna kammare. Hos dessa djur är de båda kretsloppen därför parallellkopplade, inte seriekopplade. Därmed kan djuren ha olika blodflöden i de båda kretsloppen. Grodor kan strypa blodflödet till lungorna och ändå ha ett högt flöde i kroppkretsloppet, vilket förmodligen är energibesparande. Detta gör de när lungorna inte användes under dykning, till exempel när syrgasen i dem är förbrukad eller när lungorna inte behövs. Grodor som övervintrar i botten av vattensamlingar kan andas enbart genom huden. Eftersom deras syrgasbehov är lågt i kylan, klarar de sig utan att andas via lungorna. Dykande däggdjur, som valar och sälar, brukar inte använda lungorna som syrgasförråd. Men de måste ändå pumpa samma volym blod per minut till lungkretsloppet som de gör till kroppskretsloppet. Fördel till grodorna, alltså.

Det står ofta i skolböckerna att syrgasrikt blod blandas med syrgasfattigt i ett trerummigt hjärta. Blodet som når kroppens organ skulle alltså innehålla betydligt mindre syrgas hos djur med trerummigt hjärta, än det gör hos oss. Detta stämmer inte med verkligheten. Det blod som grodor och reptiler pumpar ut i kroppskretsloppet består till allra största delen av syrgasrikt blod från lungorna. Många reptiler har en rad fiffiga anordningar i hjärtats enda kammare som håller det syrgasrika blodflödet från lungorna separerat från det syrgasfattiga flödet från kroppens organ. Därmed kommer det syrgasrika blodet pumpas vidare till kroppens organ och det syrgasfattiga till lungorna. Även grodorna har anordningar som håller flödena skilda åt. Ingen större nackdel för grodor och ödlor, alltså.
Vad är fördelen med att ha fyra rum i hjärtat och två skilda kretslopp?
Men vad är då fördelen för oss däggdjur med att ha ett fyrrummigt hjärta och två separata kretslopp kopplade i serie? Förmodligen är fördelen att trycket kan var högt i kroppskretsloppet. Hos djur med trerummigt hjärta måste trycket vara lika i de båda kretsloppen. Det är ju samma kammare som producerar det drivande trycket för båda kretsloppen. Trycket i lungkretsloppet kan inte vara för högt. Då pressas alltför mycket vätska ut ur blodkapillärerna och lungblåsorna blir vätskefyllda, vatten i lungorna med andra ord. Hos djur med trerummigt hjärta sätts därmed en övre gräns också för trycket i kroppskretsloppet. Det kan ju inte vara högre än trycket i lungkretsloppet.
Men om du har ett fyrrummigt hjärta och två seriekopplade kretslopp, kan du ha olika tryck i de båda kretsloppet. Trycket i lungkretsloppet produceras ju av höger kammare och trycket i kroppskretsloppet av vänster. Fåglar och däggdjur har mycket höga tryck i kroppkretsloppet och ändå låga i lungkretsloppet, så att inte vatten ansamlas i lungorna. Vad är då fördelen med ett högt tryck i kroppskretsloppet? Det vet vi faktiskt inte! Ett högt tryck är inte nödvändigt för att åstadkomma ett högt blodflöde. Ett högt blodflöde kunde vi fått också genom att minska resistansen ("motståndet") i blodkärlen och då hade dessutom hjärtats energibehov blivit mindre. I ett högtryckssystem är det kanske lättare att omfördela blodet mellan olika organ, till exempel att "shunta" blod från magtarmkanalen till de arbetande musklerna vid fysiskt arbete. Men vi vet, som sagt, inte hur det förhåller sig med den saken.
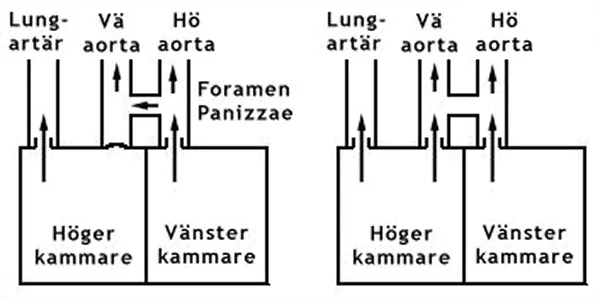
Krokodildjurens fyrrummiga hjärta
Krokodilerna då? Jo, krokodilernas cirkulation fungerar normalt som vår. Se på bilden ovan. Med sitt fyrrummiga hjärta kan de ha två separata seriekopplade kretslopp, precis som vi däggdjur. Men krokodilerna kan också, precis som grodorna, strypa blodflödet till lungkretsloppet, utan att minska flödet till kroppkretsloppet lika mycket. Från höger kammare utgår både lungartären och en av krokodilernas två aortor. Den andra aortan utgår från vänster kammare.
Under normala förhållanden pumpas allt blod från höger kammare via lungartären till lungorna. Klaffen till höger kammares aorta är stängd, eftersom blodtrycket är högre i aortorna än i höger kammare. Det finns ett hål som förbinder de båda aortorna (foramen Panizzae). Blod från vänster kammare pumpas i större eller mindre utsträckning genom detta hål till den aorta som inte tar emot blod från höger kammare. Cirkulationen blir som i den vänstra bilden ovan. Krokodilen har nu två seriekopplade kretslopp, precis som vi däggdjur. Den har också ett lägre tryck i lungkretsloppet än i kroppskretsloppet, precis som vi.
Men krokodilen kan strypa blodflödet till lungorna, bland annat genom en slutmuskel, som fungerar som en ventil i början av lungartären. Då stiger trycket i höger kammare, klaffen till höger kammares aorta öppnas och en del av höger kammares blod pumpas ut till kroppen via denna aorta. Cirkulationen blir då som i den högra bilden ovan. Krokodilen har nu två parallellkopplade kretslopp, precis som groddjur och kräldjur. Krokodilerna använder sig kanske av denna möjlighet att föra över blod från lungkretsloppet till kroppskretsloppet när de dyker eller när de håller andan länge på land. Om syreförrådet i lungorna är litet eller förbrukat, har lungkretsloppet ingen funktion. Detta är därför troligen energibesparande att pumpa en mindre mängd blod till lungorna.
En annan möjlighet är att krokodilerna pumpar ut syrefattigt blod i den ena aortan för att underlätta avsöndringen av saltsyra till magsäcken. Syrefattigt blod är nämligen också försurat av koldioxid, som ger upphov till kolsyra. Därmed har detta blod en högre halt av vätejoner. Om halten av vätejoner är hög i det blod som når magsäcken, krävs det mindre energi för att aktivt transportera in vätejoner i magsäckens hålrum. Krokodiler behöver ha en mycket sur magsaft för att kunna lösa upp de stora köttstycken och skelettben, som de sväljer. De tuggar nämligen inte födan.
Vi skulle kunna hävda att krokodilerna har det bästa hjärtat bland ryggradsdjuren. Deras hjärta tycks i varje fall kombinera fyrrummighetens fördelar med trerummighetens. Men vi har ännu inte fullständigt förstått hur krokodilhjärtat fungerar.
Krokodilerna är, näst efter fåglarna, de utdöda dinosauriernas närmaste nu levande släktingar. Fåglarna räknas numera som en nu levande undergrupp bland dinosaurierna, medan de krokodilartade reptilerna bildar en egen grupp. Hade dinosaurierna ett "krokodilhjärta" eller ett "fågelhjärta"? Forskare har hittat ett dinosauriefossil som troddes innehålla ett bevarat hjärta. Nyare studier tyder dock på att denna struktur inte är ett hjärta, utan helt av oorganiskt ursprung. Men en rimlig gissning är att de utdöda dinosaurierna hade ett fyrrummigt hjärta och samma cirkulatoriska kopplingsschema som fåglarna. Fåglarna är ju levande dinosaurier.
Fördjupning
Har myror ett hjärta? Om insekternas öppna cirkulation och människans slutna blodomlopp. Du hittar artikeln under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
Referenser
Texten har uppdaterats och utökats år 2013.
- J. Hecht: Wild at heart. Were dinosaurs pumped up for a racy lifestyle? (New Scientist 166:8, 29 april 2000).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- D. Randall, W. Burggren och K. French: Eckert's animal physiology, mechanisms and adaptations (4:e uppl, Freeman, 1997).
- K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).
- P.C. Withers: Comparative Animal Physiology (Saunders College Publishing, 1992).

Människans hjärta är inte en pump, utan två pumpar. Dess högra del pumpar ut blod till lungorna, dess vänstra del till alla kroppens organ. Båda delarna drivs av hjärtmuskulaturens sammandragningar och slår exakt samtidigt. Hur styrs deras aktivitet under loppet av ett hjärtslag?
Hjärtslaget i vila
Vid en normal vilopuls på 70 hjärtslag/minut varar ett hjärtslag 0,86 sekunder, vid en maxpuls på 200 slag/minut under intensivt arbete hos en ung person bara 0,30 sekunder. Maxpulsen minskar med åldern. Pulsen kallas också hjärtats slagfrekvens. Under en livstid på 80 år slår ett människohjärta, mycket grovt räknat, 3 miljarder slag. Denna fantastiska pump kan klara av detta utan underhåll utifrån. Men till slut är det ändå den som för många av oss begränsar det ögonblick i tiden som är ett människoliv. Hjärt- och kärlsjukdom är den vanligaste dödsorsaken i Sverige och står för cirka en tredjedel av alla dödsfall.
Elektriska impulser genom retledningssystemet
Under ett hjärtslag stimuleras hjärtats olika delar till kontraktion (sammandragning) genom att elektriska nervimpulser (aktionspotentialer) vandrar längs med de elektriskt kopplade hjärtmuskelcellernas cellmembraner. Nervimpulserna följer normalt alltid samma väg längs med det så kallade retledningssystemet. Notera "nervimpulserna" sprids via hjärtmuskelceller, inte nervceller. De uppkommer spontant i sinusknutan (sinusnoden) i höger förmak, sprids över förmaken till AV-knutan (AV-noden) mellan förmaken och kamrarna, löper i skiljeväggen mellan kamrarna till hjärtats nedre spets och sprids sedan över hela kammarmuskulaturen.
Sinusknutan fyrar av spontant, men dess frekvens kan ändras av nervstimuli och adrenalin
Sinusknutan fyrar av med en spontan inneboende frekvens på cirka 100 slag per minut. Men dess frekvens påverkas ständigt av hormonet adrenalin och av det så kallade sympatiska nervsystemet, som båda höjer frekvensen, och av det så kallade parasympatiska nervsystemet, som sänker frekvensen. I vila dominerar parasympatisk stimulering. Vilopulsen är därför lägre än 100 slag per minut. Vid fysiskt arbete dominerar sympaticus och adrenalin, vilket leder till att pulsen ökar.
De högsta och lägsta slagfrekvenserna hos däggdjur och fåglar
Hos flygande fladdermöss, hårt arbetande näbbmöss och flygande kolibrier har forskare, otroligt nog, uppmätt en puls på mer än 1 000 hjärtslag per minut, svarande mot mer än cirka 0,06 sekunder per hjärtslag. Hos näbbmusen Suncus etruscus, ett av de minsta däggdjuren, har forskarna registrerat ett rekord på 1 511 slag per minut. Detta motsvarar en frekvens på 1 511 Hz. För ljud ligger denna frekvens nära mitten av talområdet. Det är en gåta hur näbbmusens hjärta kan hinna med att fyllas och pumpa ut blod på avsevärt mindre än en tiondels sekund. Å andra sidan har forskare hos gnagare i vinterdvala och hos dykande sälar uppmätt färre än 10 hjärtslag per minut, tidvis färre än 5.

Elektrokardiogrammet (EKG)
Ett elektrokardiogram (EKG) återspeglar hela hjärtats elektriska aktivitet. Aktionpotentalernas färd genom hjärtat leder till att en elektrisk spänning uppkommer över hela hjärtat. Hela hjärtat fungerar alltså som ett elektriskt batteri. Batteriets spänning och det elektriska fältets riktning varierar på ett lagbundet sätt under loppet av ett hjärtslag. Om vi ansluter elektroder till lämpliga ställen på kroppsytan, kan vi med hjälp av en EKG-apparat mäta hur spänningen förändras med tiden. I EKG:et överst på denna sida kan du se fyra fullständiga hjärtslag.

Systole, diastole och förmakskontraktion
På figuren ovan ses ett schematiskt EKG med beteckningar för topparna och dalarna. I figuren kan du också se hur EKG:et återspeglar kontraktionen (sammandragningen) och relaxationen (avslappningen) av hjärtats olika delar. Under systole kontraheras de båda kamrarna och pumpar ut blod i aortan och lungartären. Under diastole relaxeras kamrarna och fylls med blod. Under den korta förmakskontraktionen& kontraheras de båda förmaken och pumpar in blod i kamrarna. Förmaken står, konstigt nog, bara för en liten del av kamrarnas fyllning. Kamrarna fylls till största delen innan förmakskontraktionen, genom att blod flödar genom de relaxerade förmaken. Detta sker dels genom att ventrycket pressar blod in i kamrarna, dels genom att kamrarna utvidgas som elastiska gummibollar och suger in blod från venerna.
Referenser
- K. E. Barnett, S. M. Barman, S. Boitano, och H .L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal physiology (3rd ed, Sinauer, 2012 ).
- K. D. Jürgens et al.: Heart and respiratory rates and their significance for convective oxygen transport rates in the smallest mammal, the Etruscan shrew Suncus etruscus& (Journal of Experimental Biology 199:2579-2584, 1996).
- B. I. McDonald och P. J. Ponganis: Deep-diving sea lions exhibit extreme bradycardia in longduration dives (Journal of Experimental Biology 217:1525-1534, 2014).
- W. K. Milsom, M. B. Zimmer, och M. B. Harris: Regulation of cardiac rhythm in hibernating mammals (Comparative Biochemistry and Physiology Part A 124:383-391, 1999).
- R. K. Suarez: Hummingbird flight: sustaining the highest mass-specific metabolic rates among vertebrates (Experientia 48:565-570, 1992).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).

För de flesta djur är syrebehovet deras mest akuta behov. Därnäst kommer behovet att avge koldioxid, som bildas i kroppen vid cellandningen. Cellandningen producerar energi i form av molekylen ATP, som driver alla kroppens energikrävande processer. Mat kan många djur klara sig utan i dagar eller veckor. Och vatten kan många landdjur undvara i timmar eller dagar. Men syrebrist leder ofta till problem inom sekunder eller minuter. Det finns naturligtvis många undantag, djur som kan leva mycket länge utan syre och som tål ansamling av koldioxid i kroppen. Denna artikel ska handla om hur djur tar upp det nödvändiga syret från omgivningen och avger den koldioxid, som bildas i kroppen.
Ser vi utvecklingsmässigt på det hela så andades de första djuren genom huden. När djuren blev mer komplexa uppkom gälar. Men hos olika djurgrupper uppkom gälarna på olika ställen, till exempel i främre delen av tarmen hos ryggradsdjur och som utskott från benen hos många kräftdjur. Alla gälar har alltså inte samma evolutionära ursprung. När djuren kom upp på land utvecklades nya andningsorgan: lungor och trakéer. Men luftandningsorganen utvecklades som regel inte ur gälapparaten. Lungorna hos ryggradsdjuren utvecklades som en utbuktning från tarmen och trakésystemet hos leddjur som inbuktningar från huden. Så även för luftandningsorganen gäller att de har olika evolutionärt ursprung hos olika djurgrupper.
Små djur behöver inte några andningsorgan. De får tillräckligt med syre genom huden via diffusion.

En del groddjur har förlorat lungorna och andas bara via huden och munhålans väggar. Till dem räknas alla arter inom den mycket artrika familjen Plethodontidae, två andra salamanderarter, en art bland de benlösa maskgroddjuren och en nyupptäckt grodart på Borneo. Groddjurens hud är, till skillnad från andra fyrfota ryggradsdjurs, fuktig och genomtränglig för vatten, syre och koldioxid. De flesta groddjur andas faktiskt inte bara med lungorna, utan också med huden och munhålan.
Två transportmekanismer
Två transportmekanismer behövs i andningsorgan: diffusion, som drivs av en koncentrationsskillnad, och massflöde, som drivs av en tryckskillnad. Diffusion är bara effektiv över korta avstånd, mindre än cirka 1 millimeter i vatten, dock längre sträckor i luft. Massflöde kan däremot transportera syre och koldioxid över mycket långa avstånd i både andningsorgan och i blodomlopp.
Tre typer av andningsorgan
Större djur måste ha andningsorgan. Ett andningsorgan är i princip en ytförstorad del av djurets kroppsyta. Ytförstoringen gör att gasutbytet med hjälp av diffusion blir effektivare. Diffusion sker dock bara den sista korta delen av transportvägen från omgivningen till kroppens inre, över ett eller ett par tunna cellskikt från vatten eller luft till blodet eller till en annan kroppsvätska. Över längre avstånd är diffusionen inte tillräckligt effektiv. Därför finns det för det mesta en mekanism genom vilken mediet utanför djuret (det vill säga vattnet eller luften) transporteras till och från andningsytan genom massflöde. Denna transport kallas ventilation. Som nämnts ovan, finns det tre huvudtyper av andningsorgan: gälar, trakéer och lungor.
Gälar
Gälar är ytförstorade utbuktningar från kroppsytan som för det mesta används i vatten. De kan vara flikiga, fjäderlikt förgrenade eller försedda med en mängd tunna skivor (lameller).
Yttre gälar finns på kroppsytan. Yttre gälar kan hos stillasittande djur ventileras antingen passivt av vattenströmmar eller aktivt genom att djuret viftar med gälarna, som om de var solfjädrar. En nackdel i det första fallet är att djuret kan drabbas av syrebrist om strömmarna upphör. En nackdel i det andra fallet är att ventilationen kan bli mycket energikrävande, åtminstone för större djur. Yttre gälar kan också ventileras aktivt av vattnet, som strömmar förbi dem, när djuret rör sig. Nackdelen med det är vattenmotståndet blir stort, vilket gör att det kostar mer energi att simma, särskilt vid höga hastigheter. På grund av dessa nackdelar har de flesta gälandande djur inre gälar.

Inre gälar är inneslutna i en gälkammare. Vattnet pumpas genom gälkammaren och flödet är enkelriktat. Gälkammaren har således både en ingång och en utgång. Ventilationen kan, som hos musslor, ske med hjälp av cilier (flimmerhår). Men oftast ventileras gälarna, som hos fiskar, av muskeldrivna pumpar.

Trakéer
Trakéer finns hos insekter och en del andra leddjur. Trakéer används bara för luftandning. Trakésystemet är ett starkt förgrenat system av luftfyllda rör. De finaste rören kallas trakeoler och är lika tunna som blodkapillärer. Trakeolerna slutar blint omedelbart intill vävnadernas celler. Hos mindre insekter sker transporten av syre genom diffusion från omgivningen, genom trakéerna och in i vävnadscellerna. Diffusionen har nämligen mycket längre räckvidd i luft än i vatten, eftersom luft är mer "lättflytande". Hos större insekter ventileras de yttre delarna av trakésystemet med hjälp av pumpande rörelser med kroppen, men i de inre delarna transporteras syret också hos dem med diffusion. Eftersom de har sitt trakésystem, behöver insekter inte transportera syre och koldioxid med blodet. Trakéer är mycket effektiva. Flygmusklerna hos vissa insekter har den högsta syrekonsumtion som över huvud taget uppmätts i hela djurvärlden. Men kanske är det trakésystemet som gör att insekter måste vara små djur.

Lungor
Lungor finns nästan bara hos luftandande djur. Lungor är ytförstorade inbuktningar från kroppsytan. Lungor kan vara säckar vars inre yta förstoras av veck eller tvärväggar (till exempel hos grodor och reptiler). Lungor kan också vara ett förgrenat rörsystem där de minsta rören slutar blint med små säckar som kallas alveoler (hos däggdjur). Andningsytan blir då stor genom att det finns väldigt många alveoler. I alla lungor överförs syret från lungluften till blodet. Fåglarna har en komplicerad andningsapparat försedd med ett antal luftsäckar.
Lungor har en så kallad tidal ventilation. Det innebär att frisk luft tas in i lungan varefter gasutbytet sker. Sedan transporteras luften tillbaka ut ur djuret samma väg. En tidal ventilation gör att vattenförlusterna vid andningen blir mindre.

Varför använder landdjur inte gälar?
Hur hade det gått om vi människor hade haft enkelriktad ventilation som i fiskarnas gälar? Då hade vi tagit in luften via munnen och släppt ut den genom ett hål på ryggen. Men vattenförlusterna hade blivit alldeles för stora. Därför ser du inga landdjur som är konstruerade på det viset. Annars hade det varit bra att bara öppna munnen och låta fartvinden ventilera lungorna under joggningsturen. Precis så gör många fiskar. De låter "fartströmmen" ventilera gälarna när de simmar snabbt. Ett fåtal landdjur, till exempel vissa krabbor, använder dock gälar på land. Men de är dåligt anpassade till att leva i luft.
En viktig anledning till att nästan inga landdjur har gälar med enkelriktad ventilation är alltså att vattenförlusterna hade blivit för stora. Dessutom skulle gälarna ha fallit ihop på land genom tyngdkraften och vattnets ytspänning. Det är därför fiskar drabbas av syrebrist när de kommer upp på land: gälarna klibbar ihop och diffusionsytan och diffusionsavståndet blir drastiskt reducerade. Så fiskarna har ingen nytta av att luften faktiskt är mycket syrerikare än vatten.
Fördjupning
- Hur sker transporter i djurens kroppar? Diffusion och massflöde. Du hittar artikeln under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
- Varför är insekter så små? Varför är däggdjur och fåglar stora? Artikeln finns under rubriken Varför är vissa djur stora medan andra är små?.
- Varför har vi två näsborrar, men en luftstrupe? Om luktsinne, andning och livet på land. Du läser artikeln under rubriken Hur utvecklades olika kroppsdelar och organ? Och hur fungerar de?
Referenser
Texten har uppdaterats och utökats år 2016.
- D. Bickford, D. Iskandar och A. Barlian: A lungless frog discovered on Borneo (Current Biology 18:R374-R375, 2008).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).
- P.C. Withers: Comparative Animal Physiology (Saunders College Publishing, 1992).

Hos alla landlevande djur påverkas blodomloppet och blodtrycket av tyngdkraften. Effekten blir särskilt stor hos stora djur med lodrät kroppställning, i synnerhet om de har långa ben och lång hals. Människans förfäder började att gå på två ben och att hålla kroppen lodrät så sent som för några miljoner år sedan. Vi är ännu inte perfekt anpassade till vår upprätta kroppsställning.
Upprätt kropp, långa ben och lång hals påverkar blodtrycket
De cirkulatoriska styrmekanismer som reglerar blodtrycket kan fallera i upprätt kroppställning även hos friska individer, i synnerhet om de är unga och långa. Om du reser sig upp snabbt, kan du drabbas av blodtrycksfall, som försämrar hjärnans syretillförsel och leder till yrsel. Om du står stilla länge kan blod och vävnadsvätska ansamlas i benen, vilket kan sänka artärernas blodtryck och orsaka svimnning.
I det flesta fall motverkar emellertid cirkulatoriska reflexer effekten av att du reser sig upp. Hjärtats slagfrekvens (pulsen) ökar. De minsta blodkärlen på blodomloppets artärsida (arteriolerna) drar ihop sig, varvid deras diameter minskar och kärlsystemets resistans ("motstånd mot flöde") ökar. Båda dessa reflexer höjer blodtrycket i artärerna. Dessutom kan venerna i nedre delen av kroppen dra ihop sig, så att deras volym minskar. Då förflyttas blod från den nedre till den övre delen av kroppen.
Det är inte bara människor som kan ha problem med tyngdkraften. Många hovdjur har långa ben. Giraffen och strutsen har både lång hals och långa ben. Alla dessa djur har ett stort lodrätt avstånd mellan huvudet och fötterna. Samma sak gäller naturligtvis för en orm med kroppen i lodrät ställning. Detta ställer till problem för blodcirkulationen, eftersom effekterna av tyngdkraften blir stora. Vi skulle kunna tro att det beror på att det blir svårt att pumpa upp artärblodet från hjärtat till hjärnan och att återföra venblodet från nedre delen av kroppen till hjärtat. Men så enkelt är det inte. Förenklat kan vi säga att effekten av tyngdkraften på artärsidan och vensidan tar ut varandra. Mera korrekt uttryckt, så handlar det om att en minskad lägesenergi i en vätskepelare helt kompenseras av en ökad tryckenergi och vice versa. Hade kärlsystemet bestått av metallrör som inte läckte, så hade tyngdkraften inte påverkat cirkulationen alls. Problemen med tyngdkraften orsakas av att blodkärlssystem är uttänjbara och av att de läcker.
Tyngdkraftens effekter på blodomloppet
Varför svimmar då en människa som stått stilla länge? Det beror på ett ökat blodtryck i nedre delen av kroppen. Tyngdkraften påverkar de blodpelare som går från hjärtat till fötterna och från hjärtat till huvudet. Den ger upphov ett så kallat gravitationstryck (hydrostatiskt tryck). Gravitationstrycket under hjärtat är positivt, vilket innebär att det är högre än atmosfärens tryck och det ökar dessutom i riktning mot fötterna. Gravitationstrycket ovanför hjärtat är negativt, vilket innebär att det är lägre än atmosfärens tryck och blir allt lägre, ju närmare hjässan du kommer. Gravitationstrycket på hjärtats nivå är således lika med atmosfärstrycket.
Låt oss förklara begreppet gravitationstryck. Vi antar att lufttrycket är 1 atmosfär. När du dyker ner under vattnet så ökar gravitationstrycket med ungefär 1 atmosfär för var tionde meter, något som du kan känna av i öronen. På cirka 10 meters djup är således trycket 2 atmosfärer, alltså dubbelt så högt som lufttrycket vid vattenytan. Om det är en meter från hjärtat till fötterna så har du i upprätt ställning ett gravitationstryck i fötternas blodkärl som är cirka en tiondels atmosfär högre än det omgivande lufttrycket. Detta tryck adderas till det tryck som hjärtats muskelkontraktioner skapar i blodkärlen. Notera att det bara är det av hjärtat skapade trycket, som driver blodet genom artärerna, kapillärerna och sedan via venerna tillbaka till hjärtat. Som nämnts ovan, är ju effekten av gravitationstrycket lika stort på artärsidan, där blod förs från hjärtat till fötterna, som på vensidan, där blod förs från fötterna till hjärtat. Gravitationstrycket motverkar således inte direkt blodflödet. Men det har indirekta effekter.
Det höga totala trycket i benen, det vill säga hjärtrycket plus gravitationstrycket, påverkar vener och kapillärer. Effekterna märks om du står stilla länge. Då utvidgas venerna i benen av det höga trycket. Blod ansamlas således i nedre delen av kroppen. Effekten av detta blir i princip densamma som om du lidit en blodförlust, nämligen minskad blodvolym och sänkt blodtryck i den övre delen av kroppen, där hjärtat och hjärnan är belägna. När du står stilla länge leder dessutom det höga kapillärtrycket i benen till att mer vätska än normalt pressas ut ur kapillärerna till den omgivande vävnaden. Blodvolymen minskar då. Även detta ger en effekt som motsvarar en blodförlust. Precis som efter en stor blodförlust kan det arteriella blodtrycket i övre delen av kroppen sjunka till den grad att hjärnan får för lite blod. Du svimmar då av på grund av syrgasbrist i hjärnan. Detta drabbar ibland soldater, som står i givakt länge. Svimningen är faktiskt funktionell, eftersom effekterna av tyngdkraften elimineras när kroppen hamnar i vågrätt läge. Även om du inte svimmar, så sväller benen och fötterna upp av den vätska som pressas ut ur kapillärerna. Sådan ansamling av vätska utanför blodkärlen kallas ödem. Notera än en gång att gravitationstrycket inte direkt motverkar blodflödet, men väl indirekt.

Gravitationens effekter på människans blodcirkulation
För den intresserade ges här en utförlig förklaring av blodtrycken och tyngdkraftens effekter på cirkulationen. Bilden visar förenklat de arteriella och de venösa blodtrycken i hjärnan, i nivå med hjärtat och i fötterna hos en upprätt 180 cm lång människa. Siffrorna är ungefärliga, men realistiska. Trycken anges i (mm Hg). Ventrycken är mycket lägre än artärtrycken. Detta beror på att resistansen ("motståndet mot blodflöde") är mycket hög i kapillärsystemens tunna kärl. Därför går en mycket stor del av den av hjärtat producerade tryckenergin förlorad, när blodet flödar från artärer till vener.
Trycken anges dels som absoluta tryck, motsvarande de tryck vi mäter med en barometer (extra fet stil), dels i relativa tryck (mindre fet stil). Inom fysiologi och medicin anges alltid relativa tryck. Detta innebär att vi använder atmosfärstrycket som referens och sätter detta tryck till 0 (mm Hg). Antag att artärtrycket mäts till 120/70 (mm Hg) och att atmosfärstrycket är 760 (mm Hg). Detta innebär att det högsta artärtrycket är 120 (mm Hg) högre än atmosfärstrycket, vilket motsvarar ett absolut tryck på 880 (mm Hg). Det lägsta artärtrycket är 70 (mm Hg) högre än atmosfärstrycket, vilket motsvarar ett absolut tryck på 830 (mm Hg). Det högsta (systoliska) artärtrycket mäts under under hjärtats sammandragningsfas. Det lägsta (diastoliska) artärtrycket mäts under under hjärtats avslappningsfas. På bilden anges medelartärtrycket, det vill säga medelvärdet av trycken under hjärtats hela arbetscykel, samt ventrycket, som varierar mycket lite under arbetscykeln.
Trycket i cirkulationen är i upprätt ställning lika med summan av det tryck som hjärtat skapar och det tryck som är en effekt av tyngdkraften, gravitationstrycket. Om du ligger ner i vågrätt läge, kan du försumma gravitationstrycket, men inte när du står upp. När vi står stilla i upprätt läge, har vi däremot en lodrät arteriell vätskepelare och en lodrät venös vätskepelare, som båda påverkas av tyngdkraften. I båda pelarna är trycket på hjärtnivå atmosfäriskt, alltså 0 (mm Hg) mätt som relativt tryck. De båda pelarna är förbundna med varandra i ändarna. Hela sytemet är således slutet. När vi förflyttar oss neråt i pelarna, stiger gravitationstrycket, precis som när vi simmar neråt under ett dyk, och blir högre än atmosfärstrycket. Under hjärtat kan pelarna jämföras med ett u-rör. När vi förflyttar oss uppåt från hjärtat i vätskepelarna, sjunker gravitationstrycket, precis som när vi simmar uppåt under ett dyk. Det blir till och med blir lägre än atmosfärstrycket, alltså negativt relativt detta tryck. De båda pelarna ovanför hjärtat kan jämföras med ett upp- och nedvänt u-rör.
När vi dyker i vatten är gravitationstrycket atmosfäriskt vid vattenytan, överst i vätskepelaren. Vi kan då fråga sig varför det inte är så i blodkärlssystemet. Där är ju gravitationtrycket atmosfäriskt på hjärtats nivå. Förklaringen till detta är komplicerad. Men det beror dels på att kärlsystemet är elastiskt och uttänjbart, dels på att de mekanismer som styr blodtrycket försvarar det totala blodtrycket på hjärtnivå, när vi reser oss upp.
Summerar vi det tryck hjärtat skapar med gravitationstrycket, finner vi att de totala trycken i artärer och vener är avsevärt högre i fötterna än på hjärtnivå. Därmed blir även trycket högre i de mellan artärerna och venerna belägna kapillärerna.
I hjärnan är däremot de totala trycken i artärer och vener är avsevärt lägre än på hjärtnivå. Det totala ventrycket är till och med negativt, alltså lägre än atmosfärstrycket. Varför kollapsar då inte huvudets vener, hoppressade av atmosfärstrycket? För hjärnans vener är detta inget problem, eftersom de med hjälp av hjärnvävnaden är "upphängda" i skallens väggar. Hjärnvävnaden är i princip en vätska och vätskors volym påverkas nästan inte alls av tryckändringar. Venerna i den övre delen av halsen tenderar däremot att pressas ihop helt av atmosfärstrycket utanför dem, när artärtrycket är som lägst.
Om kärlen ovanför hjärtat hade varit stela slangar med täta väggar, hade cirkulationen där kunnat fungera som en hävert. Om du fyller en slang med vatten utan luftbubblor, placerar den ena änden i en vattenbalja och den andra utanför baljan och under baljans vattenyta, så kan du tömma baljan. Vattnet strömmar då först uppåt i slangen och sedan nedåt. I slangens slynga kompenseras ändringarna i lägesenergi helt av ändringarna tryckenergi. Därför kan vattnet rinna uppåt i början på slangen. Orsaken till att baljan töms är att summan av tryckenergin och lägesenergin är högre i baljan än i slutet på slangen utanför baljan. Tyvärr används hävertmetoden också av kriminellt disponerade personer, när de stjäl bensin ut bensintankar.
Ökat venöst återflöde och venklaffar som försvar
Om du rör på sig motverkar du effekterna av tyngdkraften. Då pressas benens vener ihop av muskelsammandragningarna. Blodet i dem pumpas då mot hjärtat och den venösa vätskepelaren blir dessutom kortare. Klaffar i venerna hindrar blodet från att flyta åt fel håll. En sådan klaff består vanligen av två fickor. Fickorna fylls och täpper igen venen då blodet rör sig åt fel håll. Hos människan finns venklaffar i armarnas och benens vener, men inte i bålens.
Hur reagerar piloter och astronauter på g-krafter?
När astronauter eller piloter vid acceleration utsätts för mycket stora g-krafter i riktning mot benen, ger det precis samma effekt som om jordens tyngdkraft hade ökat. Då stiger det totala blodtrycket i benen alltför mycket. Blod och vävnadsvätska ansamlas därför i benen till den grad att blodtrycket i övre delen av kroppen blir katastrofalt lågt. Blodflödet och därmed syretillförseln till hjärnan kan då försämras så mycket, att personerna blir medvetslösa av på grund av syrgasbrist i hjärnan. Det svartnar då först för ögonen, så kallad "black out". Du kan till stor del eliminera denna effekt med hjälp av en stel tryckdräkt, som gör att den nedre delen av kroppen inte kan öka sin volym. Därmed förhindras ansamling av blod i venerna och utpressning av blod ur kapillärerna.
Om accelerationen sker i motsatt riktning, alltså med g-krafterna riktade mot huvudet, stiger blodtrycket i hjärnan och därmed trycket i hela hjärnvävnaden. Hjärnan är ju förpackad inne i den stela skallen och kan därför inte öka sin volym. Detta leder först till huvudvärk, sedan till mental påverkan. Synen försämras och synfältet blir rött av blod i näthinnan, så kallad "red out". Så småningom stryps blodflödet från hjärnan, vilket leder till syrebrist och medvetslöshet. Nu hjälper ingen tryckdräkt.
Den kunskapsrike astronauten står därför inte upprätt i sin rymdraket, blickande mot stjärnorna. Inte heller står han på huvudet. Han ligger på rygg, vinkelrätt mot färdriktningen. Detta läge ger minsta möjliga påverkan av g-kraften.
Tyngdkraftens effekter på ormar
Så lite grand om tyngdkraftens effekter på andra djur än människan. Här övergår vi till ormar.
Vattenlevande ormar har inga som helst problem med tyngdkraften. De kan hålla kroppen helt lodrät utan att blod ansamlas i svansen eller i huvudet. Tryckökningen i blodpelaren balanseras av en lika stor tryckökning i vattnet utanför ormen. Vatten är den perfekta tryckdräkten. Ormar som lever på marken har små problem med tyngdkraften, eftersom de för det mesta håller kroppen vågrät. Avståndet från kroppens högsta till dess lägsta punkt blir då mycket litet. Trädlevande ormar upplever däremot stora effekter av tyngdkraften. De måste kunna hålla kroppen lodrät, både med huvudet nedåt och med huvudet uppåt.

Hos trädlevande ormar finns en rad anpassningar som motverkar tyngdkraftens effekter på cirkulationen. Ormarna kan var korta, vilket minskar tyngdkraftens effekter när de håller kroppen lodrät. De är ofta smala med en seg kroppsvägg som inte går att töja ut. Därmed förhindras ansamling av blod i venerna och utpressning av blod ur kapillärerna. Ormarna har helt enkelt en inbyggd tryckdräkt. De har kanske också relativt täta kapillärväggar som motverkar utpressning av vätska från blodet. De kan ha högt blodtryck och hjärtat beläget nära huvudet, vilket minskar risken för att hjärnan får för litet blodflöde. De har också välutvecklade cirkulatoriska reflexer som försvarar det arteriella blodtrycket. De kan också utföra kropprörelser, varvid musklerna trycker ihop venerna och därmed förbättrar det venösa återflödet av blod till hjärtat. Märkligt nog tycks ormar sakna venklaffar.
Trädlevande ormar klarar således av att hålla kroppen lodrät, liksom vattenlevande ormar i vattnet. Vattenlevande ormar på land och marklevande ormar har det däremot värre. Forskare har bland annat gjort försök med afrikanska marklevande ormar tillörande huggormsfamiljen (inkluderande puffadder och gabonhuggorm). Ormarna placerades i rör som lutades i olika vinklar mot lodlinjen under en period av upp till 3 minuter. Märkligt nog fann de att blodflödet i karotidartären (som går till huvudet) helt upphörde redan när ormarna lutade 30–45 grader mot horisontalplanet med huvudet uppåt. När detta hände avbröt de försöken. Men dessa ormar skulle med stor sannolikhet svimma om forskarna höll dem länge lodrätt med huvudet uppåt. Ormarna klarade lutning med huvudet nedåt utan stora förändringar i karotidblodflödet, men de var mycket oroliga och försöken avbröts när de blev alltför stressade. Oron berodde kanske på högt tryck inuti kraniet som påverkade hjärnan. Troligen skulle ormarna svimmat av om försöken fortsatt, men det vet vi naturligtvis inte.
Referenser
- H. S. Badeer: Anatomical position of heart in snakes with vertical orientation: A new hypothesis (Comparative Biochemistry and Physiology 119A:403-405, 1998).
- U. I. Balldin: Acceleration effects on fighter pilots (pp. 1014-1017; I: K. B: Pandoff och R. E. Burr, Medical Aspects of Harsh Environments, Vol 2, Walter Reed Army Medical Center Borden Institute, 2002).
- K. E. Barret et al.: Ganong's review of medical physiology (23rd ed, McGraw Hill, 2010).
- W. F. Boron och E. L. Boulpaep (ed.): Medical physiology, a cellular and molecular approach (2nd ed, Saunders, 2009).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- H. B. Lillywhite: Circulatory adaptations of snakes to gravity (American Zoologist 27:81-95, 1987).
- H. B. Lillywhite: Orthostatic intolerance of viperid snakes ((Physiological Zoology 66:1000-1014, 1993).
- H. B. L illywhite, R. E. Ballard och A. R. Hargens: Tolerance of snakes to hypergravity (Physiological Zoology 69:293-303, 1996).
- B. Wyrick och J. R. Brown: Acceleration in aviation: G-force (Federal Aviation Administration, hämtad 19 september 2016).

Gaserna syre, koldioxid och kväve förekommer inte bara i gasform i luften utan också lösta i vatten, bland annat i våra och andra djurs kroppsvätskor. I luften beter sig alla gaserna på samma sätt. Men i vatten är det annorlunda. Koldioxid är lättlösligt i vatten, syre och kväve svårlösliga. Detta får en hel rad intressanta konsekvenser. När vatten är i jämvikt med luft avseende syre, är syrets koncentration i vattnet cirka 30 gånger lägre än i luften. Syre är således en bristvara för vattenlevande djur. Den höga syretillgången på land gör att landdjur kan ha en högre energiomsättning, något som troligen underlättat utvecklingen av komplicerat byggda nervsystem och uthålliga muskler. Syrets låga vattenlöslighet är också orsaken till att vi behöver syrebindande hemoglobin i blodet. Om du ökar trycket i gasfasen, löser sig mer gas i vattnet. Detta är orsaken till att dykare kan drabbas av dykarsjuka. Men vi börjar med koldioxid och kolsyrade drycker.
Du kan åstadkomma en imponerande kaskad av läsk och skum genom att släppa tabletter av godiset Mentos i en flaska kolsyrad läskedryck. Vad som händer är att de koldioxidmolekyler, som är lösta i läsken, mycket snabbt lämnar vätskan och ger upphov till stora mängder koldioxid i gasform. Du ska absolut inte äta Mentos före, efter eller under det att du dricker kolsyrade drycker. Det är farligt och kan mycket väl leda till skador på magtarmkanalen. Enligt en uppgift spridd på nätet skulle två barn ha dött, efter att ha ätit Mentos och samtidigt druckit Coca-Cola. Detta är bara en vandringssägen. Sådana sprids blixtsnabbt på nätet. Men ingenting blir sant bara för att det upprepas många gånger. De som dumt nog försökt äta Mentos och dricka läsk samtidigt, har emellertid drabbats av kraftig gasutveckling i mag-tarmkanalen. En kaskad av vätska och gas har sprutat ut genom munnen och de har varit mycket medtagna efteråt.
Kolsyrade dryckers fysik och kemi
Men varför blir det en så explosiv reaktion mellan Mentos och kolsyrade drycker? För att svara måste jag berätta en del om hur gasmolekyler beter sig lösta i vatten, något som är av stor fysiologisk betydelse. När vi kolsyrar läsk, bringar vi vätskan i kontakt med koldioxidgas med mycket högt tryck, betydligt högre än atmosfärstrycket. En stor mängd koldioxidmolekyler kommer då att vandra ner i vätskan med så kallad diffusion, drivna av det höga gastrycket ovanför vätskan. Sedan försluter vi flaskan. Överst i flaskan, ovanför läsken, finns då koldioxidgas med mycket högt tryck.
När du öppnar en läskedrycksflaska sjunker gastrycket ovanför läsken och blir lika med atmosfärstrycket. Du hör ju hur gas pyser ut. Nu är läsken övermättad på koldioxid, det vill säga den innehåller fler koldioxidmolekyler än som "får plats" vid det låga koldioxidtryck som råder i luften. Öppnar du en flaska försiktigt, i synnerhet om den är kyld, kommer den lösta koldioxiden långsamt med hjälp av diffusion att övergå till koldioxidgas, varvid gasbubblor bildas i läsken. Till slut inträder jämvikt, utan någon nettotransport av koldioxid mellan läsk och luft. Då är läsken "avslagen".
Men det finns ett antal faktorer som kan påskynda bildningen av koldioxidgas, flera av dem välkända för den erfarne läskkonsumenten. Om du värmer läsken, minskar koldioxidens löslighet i vatten och mer gas bildas. Om du skakar om läsken, kommer mer läsk i kontakt med luften och med så kallade groddar. Groddar är små korn i läsken eller ojämnheter på flaskans insida. Koldioxid kan bindas till groddarna, vilket påskyndar bildningen av koldioxidbubblor. De ovan nämnda Mentostabletternas yta är ojämn, försedd med ett otal mycket små gropar. Ojämnheterna verkar vara utformade så att de fungerar som utomordentligt bra groddar för bubbelbildning. Detta är med stor sannolikhet en mycket viktig orsak till Mentosens spektakulära effekt. En viktig orsak är också att en Mentostablett sjunker till botten av flaskan. Då bildas koldioxidgasen längst ner i flaskan och pressar drycken uppåt mot flaskans mynning. Andra orsaker är troligen sötningsmedlet aspartam, som finns i sockerfria läskedrycker, och konserveringsmedlet kaliumbensoat, som är en vanlig livsmedelstillsats. Dessa ämnen verkar minska det energitillskott som behövs för att bubblor ska bildas.
Det höga trycket i gasutrymmet ovanför läsken kan utgöra en fara. I sällsynta fall kan flaskor explodera. Detta gäller i ännu högre grad kolsyrade viner, som champagne. Dessa viners korkar kan ju också flyga i väg som projektiler. Skador orsakade av avfyrade champagnekorkar förekommer, enligt uppgift, särskilt i Frankrike. En större gasvolym ovanför drycken ökar risken. Gasens tryckenergi är nämligen lika med volymen multiplicerat med trycket. Om det inte funnits någon gas mellan drycken och korken hade risken eliminerats. Trycket i drycken skulle fortfarande vara högt, men detta skulle inte kunna leda till någon explosion vid öppnandet. Orsaken till detta är att vätskors volym, till skillnad från gasers, knappast alls ökar när trycket sjunker. Tillverkarna av champagne och läsk säger dock att det är tekniskt omöjligt att eliminera gasutrymmet i flaskorna.
Hur uppkommer bubblor i ett vattenglas?

Alla gaser har högre löslighet i vatten, ju lägre temperaturen är. Detta innebär att vatten kan rymma fler gasmolekyler per volymenhet vid låga temperaturer, än vid höga. Gaskoncentrationerna är då höga. Det kalla vattnet i glaset har från början varit i kontakt med luft. Då har framför allt syre och kväve från luften löst sig i vattnet tills dess att diffusionsjämvikt uppnåtts. Vid jämvikt sker ingen nettotransport av gasmolekyler mellan vattnet och luften. Sedan har vattnets temperatur stigit vid uppvärmningen, varvid gasernas löslighet minskat. Vattnet blir då övermättat på syre och kväve. Det innebär att jämvikt inte längre råder. Vattnet innehåller "för mycket" syre och kväve. Tillståndet är inte längre stabilt. De lösta gasmolekylerna tenderar därför att övergå i gasform som små bubblor. Detta finns emellertid en viss tröghet i systemet, mycket beroende på att vattnets ytspänning måste övervinnas, när bubblor bildas. Bildning av gasbubblor underlättas där det finns groddar, små strukturer som kan binda gasmolekyler. Det är därför som bubblorna bildas främst vid glasets vägg, där det finns små ojämnheter eller små föroreningar, som överlevt disken. Dessa strukturer fungerar som groddar för bildning av gasbubblor.
Gasers ökade löslighet vid låga och minskade vid höga temperaturer har betydelse för djurs syretillgång. I varmt stillastående vatten uppkommer lätt syrebrist. Sådant vatten är vanligt i tropikerna.
Syretiilgången för landdjur och vattendjur
Så lämnar vi de kolsyrade dryckerna och återvänder till djurvärlden. Som ovan nämnts, gynnas landdjur av den höga syrehalten i luft, jämfört med vatten. Detta gör att de kan ha en högre energiomsättning, något som sannolikt underlättat utvecklingen av komplicerat byggda organismer. Det möjliggör också uthålligare och intensivare muskelarbete. Energikostnaden för andningen, alltså utbytet av syre och koldioxid med omgivningen, är dessutom mycket lägre på land än i vatten. Troligen har den höga syrehalten också underlättat utvecklingen av avancerade hjärnor. Mycket få vattenlevande djur har utvecklat hög kognitiv förmåga ("intelligens") och komplicerad social samverkan. Ett undantag är vissa bläckfiskar, som i beteendeförsök uppvisat förvånansvärt avancerad kognition. Men dessa havets intellektuella giganter, skulle aldrig kunna utveckla en hög kultur, lik människans. Andra orsaker till detta är deras korta livslängd och det faktum att de aldrig skulle kunna lära sig att behärska elden, som olyckligtvis släcks av vatten.
Landdjurens vattenproblem
Den största nackdelen med att leva på land är att mycket vatten förloras genom avdunstning från kroppsytan och från andningsorganen, i synnerhet i torra miljöer. Väl anpassade landdjur måste därför ha en hud som är mycket vattentät. Däggdjur vattentätar huden med hjälp av dess översta döda hornlager, som är impregnerat med vattenskyende fettämnen. Insekter vattentätar sitt yttre skelett, kutikulan, med hjälp av ett mycket vattentätt vaxlager. Däggdjuren minskar vattenförlusterna via andningen genom att återvinna vatten i näshålan, där vattenånga kondenseras till flytande vatten på de kalla näsmusslorna. Insekternas andningsorgan utgörs av ett i hela kroppen förgrenat luftfyllt rörsystem, trakéerna. Ett sådant system avger mindre vattenånga till omgivningen, än vad lungor gör. Dessutom kan trakérörens mynningar stängas, när syreförbrukningen är låg.
Det är förvånande få djurgrupper som är så väl anpassade till landliv, att de kan leva i torra miljöer och är oberoende av vatten för sin fortplantning. Egentligen är det bara insekter, spindeldjur, kräldjur inklusive fåglar och däggdjur. Landlevande ringmaskar (till exempel daggmaskar), rundmaskar, snäckor, kräftdjur (till exempel gråsuggor) och groddjur måste nästan alla leva i fuktiga miljöer. En del av dessa djur (till exempel de flesta groddjur) måste dessutom fortplanta sig i vatten.
Det är värt att påpeka att koldioxid huvudsakligen transporteras i blodplasman, vätskan utanför blodkropparna, inte inne i dem. Där transporteras koldioxiden huvudsakligen i form av vätekarbonat (HCO3-), i mindre utsträckning i form av löst koldioxid (CO2). En del koldioxid binds till hemoglobinet, men inte till bindningsstället för syre. Det finns två orsaker till att hemoglobinet inte är så viktigt för koldioxidtransporten. För det första har koldioxid mycket högre löslighet i vatten än syre. För det andra reagerar koldioxiden med vatten, varvid kolsyra bildas. Kolsyran dissocieras till vätekarbonat och vätejoner, som båda är mycket vattenlösliga.
Syrets transport i blodet
Syrets låga vattenlöslighet ger upphov till ett stort problem, när det ska transporteras i blod genom kärlsystem. Transporten är mycket ineffektiv, om blodet bara transporterar lösta syremolekyler. Koncentrationen av lösta molekyler är ju mycket låg. De flesta djur med blodkärlssystem har därför syrebindande andningspigment i blodet. Den mest kända syrebindande molekylen är det röda hemoglobinet. Hemoglobin har en mycket stor benägenhet att binda syre. Därmed rymmer blodet betydligt mer syre än de fria lösta molekylerna. Tack vare hemoglobinets stora benägenhet att binda syre blir den totala syrekoncentrationen i blodet (mätt som gram syre per liter blod) i storleksordningen 100 gånger högre än den annars skulle vara. Nästan allt detta syre är bundet till hemoglobinet. Koncentrationen av fria syremolekyler är fortfarande mycket låg.
Hjärtan är muskeldrivna pumpar. Utan hemoglobin hade människans hjärta, teoretiskt sett, behövt pumpa ut i storleksordningen 100 gånger mer blod per tidsenhet för att förse kroppen med den normala syremängden. Det finns inga muskler som klarar av att dra ihop sig tillräckligt snabbt och ofta för att klara av detta arbete. Och ett hjärta med 100 gånger större volym är en anatomisk omöjlighet.
Dykarsjuka
Gasers lösta i kroppsvätskorna ställer till problem för dykare och kan bland annat orsaka så kallad dykarsjuka ("bends", tryckfallssjuka). För att förstå dykarsjuka, måste vi först ge lite bakgrundsfakta. Ju högre trycket för en gas är i luft, ju mer gas löses i en vattenlösning som är i kontakt med luften. Gasen löses i vattnet tills diffusionsjämvikt nås. Vätskan är då mättad avseende gasen. Detta diskuterades ovan för kolsyrade drycker. I luft beter sig de olika ingående gaserna som om de andra gaserna inte existerade. Vi kan därför tala om ett partialtryck för varje gas. Torr luft innehåller 20,9 volymprocent syre och 78,1 volymprocent kväve. Om det totala lufttrycket i torr luft är 760 mm Hg (1 013 hPa eller 1 atm; atm betyder atmosfär), kan vi med hjälp av procenttalen räkna ut avrundade partialtryck för syre på 159 mm Hg eller 0,2 atm och för kväve på 594 mm Hg eller 0,8 atm. Vi använder i fortsättningen enheten atm, där 1 atm är det normala lufttrycket vid havsytans nivå. I fuktig luft, som innehåller vattenånga, kommer partialtrycken för syre och kväve att vara lägre än de ovan nämnda. Vi försummar denna effekt av vattenånga i luften.
Trycket i vatten ökar med djupet under vattenytan, närmare bestämt med cirka 1 atm för var tionde meter. Vattentrycket är således 2 atm på 10 meters djup, 3 atm på 20 meters djup, 4 atm på 30 meters djup och så vidare. Äntligen är det så dags för dykaren att göra entré. Kroppsvätskorna är vid början av ett dyk i jämvikt med luft med partialtrycken 0,8 atm för kväve och 0,2 atm för syre. En dykare, som andas luft från gastuber, kommer på tjugo meters djup andas in luft med samma totaltryck som det omgivande vattnets, det vill säga 3 atm. I inandningsluften har partialtrycket för kväve då tredubblats till 2,4 atm och för syre till 0,6 atm. Nu inträder samma situation som då vi kolsyrar läskedrycker. Kväve och syre kommer att transporteras från lungorna via blodet till alla kroppens vävnader och koncentrationerna av fria gasmolekyler stiger i vävnadsvätskorna. De koncentrationer av gaserna som nås beror dels på djupet under vattenytan, dels på den dyktidens längd.

Om dykaren snabbt dyker upp till vattenytan, inträder samma situation som då vi öppnar en läskedryck. Kroppsvätskorna är övermättade på kväve och syre. Dykaren har inte hunnit andas ut överskotten av gaserna. Då bildas små gasbubblor i vävnaderna. Kvävet ger de största problemen, eftersom syrehalten är lägre än kvävehalten och syret dessutom förbrukas av kroppen. Detta är dykarsjuka, som orsakas både av bubblorna i sig och av processer som bubblorna orsakar i kroppen. Symptomen kan i lindriga fall inskränka sig till missfärgad hud, klåda och ledsmärtor. I allvarliga fall drabbas dykaren av andningsproblem, yrsel, förlamning och medvetslöshet, i värsta fall döden.
Dykarsjuka förebygges genom att dykare måste stiga upp stegvis, med pauser på olika djup. Då kan överskottet på kväve andas ut och bubblor bildas inte i kroppen. Schemat är anpassat efter dykdjupet och dyktiden. Om dykarsjuka uppkommit, får dykaren vistas i en tryckkammare med högre lufttryck än i atmosfären. Då löser sig bubblorna vanligen i kroppsvätskorna. Sedan sänker man gradvis trycket i kammaren, så att kvävet kan andas ut utan bubbelbildning. Vid långvariga dyk till stora djup kan kvävet i gastuberna ersättas med helium, som lämnar vävnaderna snabbare än kväve.
Ett annat av de många problem, som måste undvikas vid dykning, är kväveberusning (djupberusning) med därav följande omdömeslöshet. Vid höga koncentrationer har kväve (N2) nämligen liknande effekter som lustgas (dikväveoxid, N2O).
Gasers löslighet och Henrys lag
För den intresserade ges här en fördjupad förklaring till de processer som diskuteras ovan. En gas kallad G finns dels i luften ovanför en vattenlösning, dels löst i vattenlösningen. Mellan luft och lösning är det diffusionsjämvikt avseende gasen G. Det innebär att det finns ett utbyte av gasmolekyler mellan luft och lösning, men ingen nettotransport i endera riktningen (se pilarna). Lösningen är då mättad på gasmolekyler. Då gäller Henrys lag:

Förklaring av bilden:
- Prickarna symboliserar gasmolekyler.
- PG är gasens partialtryck (mm Hg eller hPa) i luften. Ökar trycket i luften kommer mer gas från luften att lösas i vattnet. Detta händer när vi kolsyrar läsk och när en dykare dyker neråt. Minskar trycket i luften blir vattnet övermättat på gasen och gasbubblor bildas. Detta händer när vi öppnar en läskflaska och när en dykare dyker uppåt för snabbt.
- CG är gasens koncentration (gram/liter) i vattnet. Den ökar, när gastrycket i luften ökar, och minskar, när gastrycket där minskar.
- αG är gasens löslighet (löslighetskoefficient) i vatten. Den är mycket låg för syre och kväve, mycket hög för koldioxid. Det är därför som hemoglobin är nödvändigt för blodets syretranport, men mindre viktig för dess koldioxidtransport.
- För alla gaserna gäller att αG ökar vid minskad temperatur och minskar vid ökad temperatur. Det är därför som läsken blir övermättad och bildar koldioxidbubblor vid uppvärmning. Det är också därför som djur i kalla vatten har tillgång till mer syre än djur i varma vatten.
I den schematiska bilden nedan är partialtrycket, PG, i luften ovanför vattnet lika stor för alla de tre gaserna. Temperaturen är också lika stor. Det sker inte heller någon nettodiffusion mellan luften och vattnet. Diffusionsjämvikt råder således i alla tre fallen. Du ser skillnaderna i CG mellan gaserna. Koncentrationen av den vattenlösligare koldioxiden är ungefär lika stor i luften som i vattnet. Koncentrationerna av de dåligt vattenlösliga gaserna syre och kväve är däremot mycket lägre i vattnet än i luften.

När jämvikt avseende en gas råder mellan luft och en vattenlösning, säger vi att gasens partialtryck råder också i vattenlösningen. Detta är en egendomlig terminologi, eftersom gasen inte utövar något gastryck i vattenlösningen. Men den är nödvändig för att kunna beskriva gasers diffusion i vatten och mellan vatten och luft.
Fördjupning
Hur sker transporter i djurens kroppar? Diffusion och massflöde. Du hittar artiklen längre ner här under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten
Varför skadas däggdjur och fåglar inte av dykarsjuka (tryckfallssjuka) vid dyk till stora djup?. Du hittar artiklen längre ner här under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten
Referenser
- K. E. Barrett, S. M. Barman, S. Boitano, och H. L. Brooks: Ganong's review of medical physiology (23rd ed, McGraw-Hill Lange, 2010).
- C. Bothin: Dykarsjuka (Internetmedicin.se, hämtad 20 juni 2016).
- C. W. Bryant: What's a diving bell? (HowStuffWorks, hämtad 21 juni 2016).
- T. S. Coffey: Diet Coke and Mentos: what is really behind this physical reaction? (American Journal of Physics 6:551-556, 2008).
- K. Dodde, B. Dodde, och I. MacQuarrie: Diving bells and observation chambers (Diving Heritage, hämtad 21 juni 2016).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- B. Mikkelson: Non compos Mentos (Snopes.com, uppdaterad 9 november 2006).
- Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).
- P. Willmer, G. Stone, och I. Johnston: Environmental physiology of animals (2nd ed, Blackwell, 2005).

Dykarsjuka (tryckfallssjuka) är ett farligt tillstånd, som kan drabba mänskliga dykare och i allvarliga fall leda till döden. Många däggdjur och fåglar dyker under längre tid och till större djup än människor. Drabbas de inte av dykarsjuka? Eller tål de dykarsjuka så att de inte påverkas av de allvarliga effekter som kan skada mänskliga dykare?
Hur uppkommer dykarsjuka?
Dykarsjuka innebär att kvävebubblor kan bildas i vävnaderna, när människor eller andra djur simmar upp till ytan efter djupdykning. Under dykning till stora djup ökar det omgivande vattnets tryck och därmed också kvävgastrycket i luften inuti lungorna. Detta leder till att extra mycket kväve från lungluften löser sig i kroppsvätskorna. Kvävekoncentrationen i kroppens vävnader blir då högre än normalt. När dykaren återvänder till ytan sjunker omgivningens tryck och därmed också kvävgastrycket i inandningsluften och lungorna. Då blir vävnaderna övermättade på kväve. Överskottskvävet återgår då till gas och bildar gasbubblor i vävnaderna. Bubblorna skadar vävnaderna och leder i värsta fall till döden. Syre ger inte upphov till dykarsjuka, eftersom det förbrukas av kroppens celler. Koldioxid ger inte upphov till dykarsjuka, eftersom det är mycket lösligare i vatten än syre och kväve.
Dykande däggdjur och fåglar fridyker
Vi vet mycket lite om hur djupdykande däggdjur, som valar och sälar undviker dykarsjuka och än mindre hur djupdykande fåglar gör det. Men det har gjorts en del studier.

Risken för dykarsjuka mindre vid fridykning än vid dykning med gastuber. I det första fallet innehåller kroppen bara det kväve som medförts när dyket inledddes. I det senare fallet tillförs ständigt nytt kväve från luften via gastuberna. Människor som dyker med gastuber undviker dykarsjuka genom att under uppstigningen göra pauser efter ett speciellt schema, innan de återvänder till ytan. Då sjunker kvävehalten i kroppen så långsamt att kvävet återförs till lungorna utan att bubblor bildas i vävnaderna.
Men även vid fridykning, i synnerhet upprepade dyk, finns det en risk för dykarsjuka. Människor har drabbats av dykarsjuka efter upprepade fridykningar. Forskare har också påvisat gasbubblor i vävnaderna hos valar och sälar, som ju rutinmässigt utför upprepade fridykningar. Det är dock oklart om gasbubblor bildas under normala dyk hos dessa djur.

Hur undviker sälar och valar dykarsjuka?
Det finns flera hypoteser som skulle kunna förklara hur valar och sälar undviker dykarsjuka. Den klassiska förklaringen är att lungorna trycks ihop, så att deras gasutbytande delar töms på luft och andningsluften hamnar i de grövre luftvägarna. De senare har en mycket liten gasutbytande yta, vilket leder till att kväveupptaget blir minimalt. Hos människor skulle bröstkorgen krossas om lungorna trycktes ihop. Hos många valar och sälar har forskarna emellertid påvisat anatomiska anpassningar som gör att bröstkorgen kan pressas ihop utan att skadas. Djuren måste emellertid dyka till rätt stora djup innan lungorna komprimeras helt. Innan de når dessa djup skulle kväve under högt tryck kunna tas upp från lungluften till vävnaderna.
Enligt en annan hypotes kan överskottskväve hos valar och sälar elimineras genom att lösas i fettvävnadernas fett för att sedan frigöras därifrån så långsamt att bubblor inte bildas. Hos två valarter har forskarna påvisat ett särskilt fett med extra hög löslighet för kväve, något som ger stöd åt denna hypotes. Hos valar finns det i huvudet ett så kallat rete bestående av en mängd parallellt löpande tunnväggiga blodkärl inbäddade i fettvävnad. Dessa kärl för syrerikt blod till hjärnan. Forskarna tror att fettvävnaden absorberar kväve och därmed skyddar nervvävnaden från bubbelbildning.
Det har också föreslagits att vävnaderna hos valar och sälar kan bli övermättade på kväve utan att gasbubblor bildas, möjligen för att de saknar så kallade groddar, små partiklar som påskyndar bubbelbildning. Ett annat förslag är att dessa djurs vävnader är anpassade till att tåla bubblor utan att skadas. Men dessa förslag har inte testats.
Hur undviker fåglar dykarsjuka?
Forskarna vet nästan ingenting om hur djupdykande fåglar undviker dykarsjuka. Det finns dock några studier av pingviner. Lungorna hos fåglar är stela. Deras storlek förändras inte under andningen. I stället pumpas luft igenom dem med hjälp av de så kallad luftsäckarna. Det har emellertid föreslagits att de vid dykning skulle fyllas med så mycket blod, att de gasutbytande delarna, luftkapillärerna, pressas ihop. Detta skulle hindra kväveupptag till vävnaderna och skydda mot dykarsjuka. Luftsäckarnas väggyta är så liten att kväveupptaget från dem troligen ät litet. Det finns dock fakta som talar emot denna hypotes.
I en intresssant studie fann forskarna att adeliepingviner och kungspingviner inte snabbt simmade rakt upp till ytan efter djupa dyk. I stället simmade de långsamt och i sned vinkel gentemot havsytan. Detta beteende fyller kanske samma funktion som mänskliga djupdykares pauser under uppstigningen, nämligen att överföra kväve till andningsluften så att gasbubblor inte bildas i vävnaderna. Flera av de andra skyddsmekanismer, som ovan diskuterats för däggdjur, skulle också kunna gälla för fåglar, men om detta vet vi intet.
Fördjupning
Artikeln Syre, koldioxid och kväve: om landliv, vattenliv, dykarsjuka (tryckfallssjuka) och kolsyrade drycker här ovanför.
Referenser
- D. P Costa: Diving physiology of marine vertebrates (Wiley Online Library, 2007).
- W. Dzik: The air they breathe (ISBT Science Series 11 Suppl. 1:317-324, 2016).
- A. Fahlman et al. To what extent might N2 limit dive performance in king penguins? (Journal of Experimental Biology 210:3344-3355, 2007).
- S. K. Hooker et al.: Deadly diving? Physiological and behavioural management of decompression stress in diving mammals (Proceedings of the Royal Society 279:1041-1050, 2012).
- P .J. Ponganis och G. L. Kooyman: Diving physiology of birds: a history of studies on polar species (Comparative Biochemistry and Physiology A 126:143-151, 2000).
- P .J. Ponganis et al.: Physiological responses of king penguins during simulated diving to 136 m depth (Journal of Experimental Biology 202:2819-2822, 1999).
- K. Sato et al.: Buoyancy and maximal diving depth in penguins: do they control inhaling air volume? (Journal of Experimental Biology 205:1189-1197, 2002).
Skulle människan kunna andas med gälar? Människan härstammar i och för sig från fiskar med gälar. Vi har också gälbågar under en del av fosterutvecklingen. Men vår kroppsbyggnad har förändrats så mycket under evolutionens gång, att det är helt omöjligt för oss att återbilda gälarna. Vi kan då fråga sig om det är möjligt att lära sig att andas vatten med våra lungor. Svaret är återigen nej. Det finns flera orsaker till att våra lungor aldrig skulle kunna fungera som organ för att andas vatten. Alla orsakerna har att göra med de stora skillnaderna mellan luft och vatten.
Lungblåsornas och de nedersta luftvägarnas byggnad

Efter att ha passerat ett stort antal förgreningar når luften de så kallade respiratoriska bronkiolerna. De saknar de broskringar som håller de grövre förgreningarna utspända. De är försedda med glatt muskulatur, på bilden återgiven som platta ljust röda band. Från de respiratoriska bronkiolerna utgår alveolgångarna. Lungblåsorna (alveolerna) är anslutna till alveolgångarna och sitter runt dem som druvorna i en druvklase sitter runt sitt skaft. Gasutbytet sker framför allt i lungblåsorna och i någon mån i de respiratoriska bronkiolerna och alveolgångarna. Små artärer (arterioler; blå) för syrefattigt blod till de respiratoriska bronkiolerna och lungblåsorna. Ett tätt kapillärnät (grönt) omslingrar alveolerna och utbyter syre och koldioxid med luften i lungblåsorna. Små vener (venoler; mörkt röda) för syrerikt blod tillbaka till hjärtat.
Respiratoriska bronkioler, alveolgångar och lungblåsor hålls utspända dels av elastisk bindväv mellan dem, dels av ett undertryck utanför dem. Undertrycket uppkommer då de elastiska lungorna drar i lungsäckarna. Lungsäckarna är tillplattade, mycket tunna vätskefyllda säckar, som är belägna mellan lungorna och brösthålans väggar. När lungorna drar i lungsäcksvätskan, sjunker trycket i den och därmed i hela brösthålan. Vätskan kan inte utvidga sig, som en gas kan. Vi kan därför säga att lungorna är upphängda i lungsäcksvätskan!
Kan vi andas vatten med lungor?
Vatten har en mycket lägre syrgashalt än luft. Vatten har dessutom en mycket högre densitet ("täthet") än luft, vilket innebär att 1 liter vatten väger enormt mycket mer än 1 liter luft. Detta innebär att vattenandande djur, för att få en given mängd syrgas, måste transportera mycket större volymer medium än luftandande djur. Den massa de behöver transportera är också mycket större. Vatten innehåller cirka 30 gånger mindre mängd syrgasmolekyler per liter än luft och ca 23 000 gånger mindre mängd syrgasmolekyler per kilogram än luft! Våra lungor är anpassade till att andas luft och vattenandning skulle ge en rad problem.
För det första är syrehalten i vatten så låg att vi inte skulle få i oss tillräckligt med syre.
För det andra hade det blivit mycket arbetsammare att andas. Våra luftvägar består av rör och de minsta rören är mycket tunna. Detta innebär att de har en relativt hög resistans ("högt motstånd") mot flöde. Det har mycket liten betydelse när vi andas luft, eftersom luften är så "lättflytande" eller, annorlunda uttryckt, den har mycket låg viskositet. Vatten är mycket mer trögflytande än luft och har alltså mycket högre viskositet. Om vi andades vatten hade därför resistansen i luftvägarna blivit mycket hög. Vi hade då behövt använda allt för stora mängder energi för att andas.
För det tredje måste vi vid lungandning transportera luften i två riktningar, först in och sedan ut. Detta spelar liten roll då vi andas luft, eftersom luften har hög syrehalt, låg densitet (se ovan) och låg viskositet (se ovan). Vid vattenandning hade förhållandena blivit radikalt annorlunda. För att få i oss samma mängd syre som vi får från ett kilogram luft, hade vi först behövt transportera 23 000 kilogram vatten in i lungorna. Där hade all rörelseenergin gått förlorad i form av värme. Sedan hade vi behövt börja på ny kula och återigen transportera de 23 000 kilona vatten ut ur lungorna. Återigen hade energiförlusterna blivit allt för stora.
För det fjärde skulle det bli problem orsakade av vattentransport med så kallad osmos. Sötvatten har mycket lägre och havsvatten mycket högre totalhalt av lösta ämnen, främst salter, än våra kroppsvätskor. Om vi andades sötvatten skulle därför vatten "sugas" in i lungornas celler med hjälp av så kallad osmos, så att cellerna till slut sprack. Om vi andades havsvatten skulle vatten "sugas" ut ur lungornas celler, så att de krympte för att till slut dö.
Experiment med vätskeandning via lungor
Forskare gjorde för ganska många år sedan försök med möss som de tvingade ner i vatten. Vattnet hade samma osmotiska egenskaper som mössens kroppsvätskor. Det hade också getts samma syrehalt som luft genom att utsättas för syrgas under högt tryck. De hade således eliminerat problemen med osmos och syrehalt. Mössen började faktiskt andas vatten. Men andningen var, av de skäl som anges ovan, mycket ansträngd. Dessutom höjdes koldioxidhalten i mössens blod successivt, bland annat på grund andningssvårigheterna. Koldioxiden gjorde att blodet försurades, vilket ledde till att mössen dog efter några minuter. Om forskarna använder vissa flytande så kallade perfluorkolväten i stället för vatten, klarar sig försöksdjuren under en längre tid. Detta beror bland annat på att syrgas har betydligt större löslighet i sådana vätskor än i vatten. Ett mål med dessa försök har varit att finna bättre behandlingsmetoder IRDS, en sjukdom som drabbar för tidigt födda barn, och ARDS, som drabbar vuxna. Båda sjukdomarna kännetecknas bland annat att lungorna blir stela, så att det blir svårt att andas in. Tyvärr har förhoppningarna att använda så kallad partiell vätskeandning vid IRDS och ARDS inte infriats.
Det har föreslagits att mänskliga dykare skulle kunna andas perfluorkolväten i stället för luft vid dyk på stora djup, bland annat för att undvika så kallad dykarsjuka. Men detta är inte ett realistiskt förslag.
Fördjupning
Läs mer i artikeln Den nödvändiga syrgasen: om djurens andningsorgan. Artikeln finns lite längre upp här, under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten
Referenser
- L. C. Clark Jr. och F. Gollan: Survival of mammals breathing organic liquids equilibrated with oxygen at atmospheric pressure (Science 152:1755-1756, 1966).
- J. B. Hancock et al.: Using liquid ventilation to improve lung function in patients with respiratory distress syndrome: a comprehensive review of the literature (AANA Journal 72:218-224, 2004).
- I. M. Galvin et al.: Partial liquid ventilation for preventing death and morbidity in adults with acute lung injury and acute respiratory distress syndrome (Cochrane Library 2013, Nummer 7).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- U. Kaisers, K. P. Kelly, och T. Busch: Liquid ventilation (British Journal of Anaesthesia 91:143-151, 2003).
- A. Kaushal, C. G. McDonnell, och M. W. Davies: Partial liquid ventilation for the prevention of mortality and morbidity in paediatric acute lung injury and acute respiratory distress syndrom (Cochrane Library 2013, Nummer 2).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).

Frågan om lungornas evolutionära ursprung har diskuterats i mer än hundra år. Det har antagits att lungorna och fiskarnas simblåsa har samma evolutionära ursprung, det vill säga att de är homologa organ, men starka belägg har saknats. Dessutom har har de båda organen olika funktioner. Simblåsor ger fiskar flytförmåga. Lungor används till att andas luft, både av fiskar och av fyrfota ryggradsdjur. Under 2000-talet har forskarna fått molekylärbiologiskt stöd för att organen verkligen är homologa. Men en fråga är fortfarande svår att besvara. Vad kom först, lungor eller simblåsa?
Molekylärbiologin tyder på samma ursprung
Forskare har jämfört transkriptomet i simblåsan hos zebrafisk med transkriptomet i lungorna hos mus och människa. Transkriptomet återspeglar vilka mRNA:n som bildas och därmed vilka gener som är aktiva. Aktiva gener producerar, via mRNA, det protein som de kodar för. Inaktiva gener producerar inte mRNA. De tre transkriptomen uppvisade stora likheter. Bland annat fann forskarna hos zebrafisken mRNA:n, som är homologa med mRNA:n för de fyra proteiner som ingår i mänskligt surfaktant. Surfaktant avsöndras in i i lungblåsorna och sänker ytspänningen i gränsen mellan vätskeskiktet på lungblåsornas insida och lungluften. Detta gör lungorna mer tänjbara och hindrar kollaps av lungblåsor.
I en annan studie har forskare visat att genen för parathyroidhormon-relaterat protein (PTHrP) är aktiv både i simblåsa och lunga. Hos alla de ryggradsdjur som undersökts är detta hormonliknande protein nödvändigt för att lungorna ska utvecklas normalt. I simblåsan påverkar det organets reaktioner på tyngdkraften. I ytterligare studier har forskare visat att en signaleringsmekanism (Wnt-transduktionsvägen) och vissa Hedgehog-gener är nödvändiga för normal utveckling i både lungor och simblåsa.

Härstammar simblåsan från lungor?
Vilket utvecklades då först, lunga eller simblåsa? Den rådande hypotesen har faktiskt varit att lungorna kom först och att de då låg på buksidan. Detta skulle innebära att förfadern till de strålfeniga fiskarna (Actinopterygii) och de köttfeniga fiskarna (Sarcopterygii) hade lungor. I så fall levde den förmodligen i sötvatten. Risken att drabbas av syrebrist kan vara stor i stillastående sötvatten. I havet är däremot nästan allt vatten mättat på syre. Nästan alla strålfeniga fiskar tillhör gruppen egentliga benfiskar (Teleostei), som utgör de flesta av de nu levande fiskarterna. Köttfeniga fiskar omfattar lungfiskar (Dipnoi) och kvastfeniga fiskar (Crossopterygii). Utdöda kvastfeniga fiskar gav upphov till alla fyrfota ryggradsdjur.
Det finns två skäl till att lungor skulle kunna vara ursprungligare än simblåsor. För det första har alla lungfiskar lungor och de enda nu levande kvastfeniga fiskarna rudimentära lungor. För det andra har tre av de fyra ursprungligaste nu levande strålfeniga fiskgrupperna lungor, nämligen fengäddsfiskar (familjen Polypteridae), bengäddor (familjen Lepisosteidae) och bågfenor (familjen Amiidae; se bild nedan). De två förstnämnda ska inte förväxlas med gäddor. Den fjärde ursprungliga gruppen, de störartade fiskarna, har simblåsa. Alla de ovan nämnda fiskarna med fungerande lungor andas också med gälar. En studie av den arteriella blodförsörjningen till lungor och simblåsor hos alla de i detta stycke nämnda fiskgrupperna stödjer tanken att de gasfyllda organen är homologa med de fyrfota ryggradsdjurens lungor.

Hos den mest avancerade strålfeniga gruppen, benfiskarna (Teleostei), antas lungorna ha förvandlats till simblåsor. De har då förlorat luftandningsfunktionen och börjat användas för att reglera flytförmågan. I samband med detta har de förflyttats från buksidan till ryggsidan. Hos en del benfiskar har simblåsan en förbindelsegång med svalget, sannolikt motsvarande luftstrupen (fysostom simblåsa). Hos andra fiskar har denna förbindelse förlorats (fysoklist simblåsa). Flera benfiskar har helt förlorat simblåsan. Hos andra har den åter igen börjat användas för luftandning.
Har lungor och simblåsa utvecklats parallellt?
Vi bör dock notera att gränsen mellan lunga och simblåsa är oklar. En lunga måste fungera som en simblåsa, eftersom den innehåller luft. Det kan vara oklart om flytförmåga eller andning är den viktigaste funktionen. Ett problem med den hittills rådande hypotesen är att lungor, utom hos bengäddor och bågfenor, ligger på buksidan, medan simblåsor ligger på ryggsidan. Den mest funktionella positionen för en simblåsa är naturligtvis ryggsidan. Detta leder till tanken att bengäddornas och bågfenornas lungor, som ligger på ryggsidan, egentligen primärt skulle fungera som simblåsor. Dessa fiskars lungor benämns också oftast simblåsor, men fysiologiskt fungerar de också som lungor.
Enligt en nyare hypotes var förfadern till de strålfeniga och köttfeniga fiskarna utrustad med ett svalg som fungerade som luftandningsorgan. I den utvecklingslinje som ledde till till de köttfeniga fiskarna, och senare till de fyrfota ryggradsdjuren, utvecklades en utbuktning på svalgets undersida till pariga lungor. Den utvecklingslinje som ledde till de strålfeniga fiskarna delades upp i två grenar. Fengäddsfiskarna (Polypteriformes) utvecklade pariga lungor på buksidan. Övriga strålfeniga fiskar, nämligen bengäddor, bågfenor, störartade fiskar och benfiskar, utvecklade en utbuktning på svalgets översida. Hos störartade fiskar och benfiskar fölorade detta utskott helt andningsfunktionen och utvecklades till en renodlad simblåsa.
Referenser
- C. Cupello et al.: Allometric growth in the extant coelacanth lung during ontogenetic development (Nature Communications 6:8222, 2015).
- M. E. Jönsson et al.: Ahr2-dependence of PCB126 effects on the swim bladder in relation to expression of CYP1 and cox-2 genes in developing zebrafish (Toxicology and Applied Pharmacology 265:166-174, 2012).
- S. Longo, M. Riccio, och A. R. McCune: Homology of lungs and gas bladders, insights from arterial vasculature (Journal of Morphology 274:687-703, 2013).
- W. K. Milsom och D. R. Jones: Characteristics of mechanoreceptors in the air-breathing organ of the holostean fish, Amia calva (Journal of Experimental Biology 117:389-399, 1985).
- S. F. Perry et al.: Which came first, the lung or the breath? (Comparative Biochemistry and Physiology Part A 129:37-47, 2001).
- S. F. Perry och M. Sander: Reconstructing the evolution of the respiratory apparatus in tetrapods (Respiratory Physiology & Neurobiology 144:125-139, 2004).
- H. Rahn et al.: Air breathing of the garfish, Lepisosteus osseus (Respiration Physiology 11:285-3, 1971).
- J. S. Torday och V. K. Rehan: Deconvoluting lung evolution using functional/comparative genomics (American Journal of Respiratory Cell and Molecular Biology 31:8-12, 2004).
- C. L. Winata et al.: Development of zebrafish swimbladder, the requirement of Hedgehog signaling in specification and organization of the three tissue layers (Developmental Biology 331:22-236, 2009).
- A. Yin, V. Korzh, och Z. Gong: Perturbation of zebrafish swimbladder development by enhancing Wnt signaling in Wif1 morphants (Biochimica et Biophysica Acta 1823:236-244, 2012).
- W. Zheng et al.: Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung (PLoS ONE 6[8]: e24019, 2011).
Människor kan inte blåsa upp sig genom att upprepade gånger pumpa in luft i lungorna, även om du kan träffa på en och annan bildligt talat uppblåst person. Grodor och paddor kan däremot blåsa upp sig, härav uttrycket "uppblåst som en groda". Förklaringen till detta är att däggdjur och groddjur andas på helt olika sätt.
Däggdjur andas in med en sugpump
Orsaken till att vi däggdjur inte kan blåsa upp oss är att vi vid inandning suger in luft i lungorna genom att utvidga dem och skapa ett undertryck i dem, så kallad undertrycksandning. Det är därför omöjligt för oss att andas in två gånger i följd utan mellanliggande utandning. Dessutom kan vi inte utvidga brösthålans och bukhålans väggar särskilt mycket. Därmed sätts en gräns för hur mycket luft vi kan ta in i lungorna. Människor kan dock något lite öka lungvolymen genom att pressa luft från munhålan och svalget ner i lungorna. Detta är dock inte ett normalt beteende, utan måste läras in.
Grodor och paddor andas in med en tryckpump
Grodor och paddor suger aldrig in luften i lungorna. De pressar alltid in den, så kallad övertrycksandning. De har faktiskt ärvt samma munhålepump (egentligen buccalpump) som fiskarna ventilerar sina gälar med. Det finns emellertid flera olika varianter hos olika stjärtlösa groddjur.

Hur grodor ventilerar sina lungor
Här beskrivs en typisk grodandning med ledning av numren på den schematiska bilden ovan. Vid (1) är grodan i utgångsläget. Vid(2)är näsborrarna öppna och glottis& (röstspringan) i struphuvudet mellan munhålan och lungorna stängd. Grodan utvidgar munhålan med hjälp av muskler (röd pil). Då sjunker trycket i munhålan så att frisk luft sugs in i den genom näsborrarna (blå pil). Vid (3) är näsborrarna stängda och glottis öppen. Grodan trycker då ihop munhålan med hjälp av muskler så att trycket i den blir högre än i lungorna (röd pil). Då pressas den friska luften in i de elastiska lungorna (blå pil).
Vid (4) är fortfarande näsborrarna stängda och glottis öppen. Munhålemusklerna har slappnat av. Eftersom trycket då blir högre i lungorna än i munhålan, pressas lungornas använda syrefattiga luft in i munhålan (blå pil). Lungorna fungerar då som vore de öppnade uppblåsta elastiska ballonger. Ofta bidrar även kroppsväggens muskler, som pressar ihop lungorna, till deras tömning. Munhålan utvidgas då (röd pil). Vid& (5) är näsborrarna öppna och glottis stängd. Munhålan trycks då ihop med hjälp av muskler (röd pil) så att den använda syrefattiga luften pressas ut genom näsborrarna (blå pil). Vid (6) är vi tillbaka i utgångsläget.
Vid (4) pressar en del groddjur utandningsluften genom munhålan och direkt ut genom näsborrarna. Då hoppar de över (5). Många groddjur pumpar mellan andetagen luft ut ur och in i munhålan, där ett visst syreupptag sker, se videon överst på denna sida.
Konsten att blåsa upp sig
Varför kan då grodor och paddor blåsa upp sig? I och med att de andas genom att skapa ett övertryck, kan de stänga glottis i struphuvudet efter en inandning, ta in ny luft i munhålan genom näsborrarna och andas in en gång till, utan mellanliggande utandning. Eftersom deras revben är korta, medan lungorna och kroppsväggen är elastiska, kan de på så sätt blåsa upp hela kroppen. Paddor gör så, om du försöker dra upp dem ur sina jordhålor, vilket leder till att de fastnar och blir svåra att avlägsna.
Strupsäckar och kväkande
Grodhanar och paddhanar har en eller två strupsäckar, som under lektiden blåses upp med hjälp av lungorna eller munhålan. I samband med detta kväker de. Strupsäckarna ökar kanske kväkandets ljudstyrka genom så kallad resonans. Alternativt gör de bara att hanen ser mer imponerande ut. En rolig spekulation är att stjärtlösa groddjur behållit den relativt ineffektiva övertrycksandningen, eftersom den gör att de kan producera mycket starka läten, nödvändiga för att honor och hanar ska träffas under lektiden.
Fördjupning
Artikeln Trerummare eller fyrarummare? Om fördelar med olika hjärtan som finns här ovanför.
Groddjur har tre olika andningsorgan
Vuxna groddjur andas inte bara genom lungorna utan också genom huden, särskilt under vatten. Dessutom fungerar, som nämnts ovan, munhålan som ett extra andningorgan i luft.
Referenser
- R. W. Hill, G .A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012 ).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- P. Willmer, G. Stone, och I. Johnston: Environmental Physiology of Animals (2nd ed, Blackwell, 2005).
Blodcirkulationen hos djur tranporterar en mängd olika ämnen till och från cellerna i kroppen. Men det som det är mest bråttom med är nästan alltid tillförseln av syre och bortförseln av koldioxid. Cirkulationssystem tenderar därför att vara dimensionerade efter djurens syrebehov. Men det finns undantag.
Insekter transporterar syre via luftfyllda trakéer
Hos insekter och en del andra leddjur ombesörjes transporten av syre och koldioxid av trakésystemet, inte av blodomloppet. Trakésystemet är ett förgrenat luftfyllt rörsystem vars finaste grenar ligger alldeles intill alla celler i kroppen, ibland till och med i inbuktningar av cellmembranet djupt inne i cellerna.
Blodkärlsystemet hos insekterna behöver alltså inte ta hand om de transporter som måste ske mycket snabbt, det vill säga tillförseln av syre och bortförseln av koldioxid. Därför klarar sig insekterna med ett relativt enkelt cirkulationssystem. Deras cirkulation är öppen, vilket innebär att de smalaste artärerna tömmer sitt innehåll i de vätskeutrymmen som finns mellan cellerna i kroppen. Kapillärer och vener saknas.
Slutna cirkulationssystem med blod
Människan och andra ryggradsdjur har ett slutet blodkärlssystem med blod. I ett sådant system rör sig blodet hela tiden inuti rörformade blodkärl, från hjärtat genom artärer, arterioler, kapillärer, venoler och vener, för att sedan flöda tillbaka till hjärtat. Arteriolerna är de smalaste kärlen på artärsidan, venolerna de smalaste på vensidan. Blodet är således skilt från vätskan utanför av kärlväggarna. Vätskan utanför kärlsystemet mellan vävnadernas celler kallas interstitialvätska. Vätskan inuti kärlsystemet mellan blodets celler (blodkropparna) kallas plasma. Plasman skiljer sig från interstitialvätskan genom att den innehåller proteiner (plasmaproteiner, till exempel albumin). Dessa proteiner kan nästan inte alls ta sig ut genom kärlväggarna och stannar därför kvar i blodet, dit de en gång har avsöndrats.
Öppna cirkulationssystem med hemolymfa
Hos insekterna, med deras öppna kärlsystem, är blodplasma och interstitialvätska samma vätska. Blodproteinerna finns både inne i och utanför kärlsystemet. Så insekter har inget blod och ingen interstitialvätska. I stället har de en enda vätska utanför cellerna som kallas hemolymfa. Hemolymfan innehåller, med mycket få undantag, inget syretranporterande pigment som vårt röda hemoglobin. Hemoglobin behövs ju inte, eftersom syre transporteras i trakésystemet. Därför finns det inte heller några röda blodkroppar. Det finns emellertid ett fåtal insekter som har hemoglobin. Öppna cirkulationssystem finns hos många ryggradslösa djur, bland annat de flesta kräftdjur och de flesta blötdjur. Bläckfiskar har dock ett slutet system.
Hemolymfan
Hemolymfan är nästan färglös och genomskinlig. Den har emellertid många viktiga transportfunktioner. Bland annat tranporterar den näring till kroppens celler, till exempel de arbetande flygmusklerna. Det handlar om sockerarten trehalos, aminosyran prolin eller fettämnet diacylglycerol, inte om glukos och fettsyror som våra celler lever på. Hemolymfan transporterar också slutprodukter från ämnesomsättningen och gifter till utsöndringsorganen, de malpighiska kärlen, som mynnar i tarmen.
Insekternas hjärta
Insekter har ett hjärta som ligger på ryggsidan, inte på buksidan som ryggradsdjurens hjärta. Se bilden nedan. Hjärtat består av ett långt rör beläget i bakkroppen. Dess vägg är försedd med muskulatur. På hjärtats båda sidor finns ett varierande antal klafförsedda öppningar, ostier (singular ostium), som enkelriktar blodflödet in i hjärtat. Ostierna är vanligen parvis ordnade med ett par i ett segment. Under hjärtat ligger en dorsaldiafragma, ett bindvävsmembran försett med hål. Ofta finns det också en ventraldiafragma på buksidan, ovanför nervsystemets längsgående bukgangliekedja. Utanför hjärtat finns det elastiska fibrer som förbinder dess yttervägg med kroppshålans vägg. Under hjärtat löper muskler mellan mellankroppens båda sidor, ibland också mellan sidorna och hjärtat. Ibland är de sammanbundna med dorsaldiafragman. De båda diafragmorna delar upp bakkroppens hålighet i tre rum: ett runt hjärtat, ett runt mag-tarmkanalen och ett runt bukgangliekedjan.
När hjärtat slår, drar muskulaturen i dess vägg ihop sig. En framåtriktad kontraktionsvåg löper då längs med hjärtat och pressar blodet framåt genom ett stort kärl som kallas aorta. Ostiernas klaffar hindrar blodet från att rinna tillbaka ut ur hjärtat. Klaffarna består av flikar som viks in i hjärtat, när det fylls, och pressas mot dess inre vägg eller mot varandra, när det drar ihop sig. När hjärtat dras samman spänns de ovan nämnda elastiska fibrerna som gummiband. När hjärtat slappnar av utvidgas det av dessa fibrer och av sin egen inneboende elasticitet. Hos vissa arter drar muskelfibrerna utanför hjärtat också ihop sig och hjälper till att utvidga det. När hjärtat utvidgas, sjunker trycket inuti det och hemolymfa sugs in i det genom ostierna.

Insekternas blodkärlssystem
Aortan löper fram genom mellankroppen till huvudet. Den drar ofta ihop sig i takt med hjärtats kontraktioner. Den sänder ut grenar till olika delar av kroppen. Grenarna tömmer hemolymfan ut i vävnaderna. Sedan sipprar hemolymfan tillbaka till bakkroppen mellan vävnadscellerna. Det finns, som sagt, inga väggförsedda kapillärer, men mellan vävnadscellerna finns det ett nätverk av hålrum. lakuner, genom vilka hemolymfan kan flyta. Det finns även större hålrum som kallas sinusar. Dessutom finns det membraner som styr hemolymfans flödesriktning. De ovan nämnda diafragmorna fyller denna funktion i bakkroppen. I benen och antennerna kan det också finnas längsgående membraner. Hemolymfan flödar då ut i benet eller antennen på ena sidan av membranet och tillbaka på andra sidan. I många fall finns små hjälphjärtan vid basen av ben, antenner eller vingar. De pumpar oberoende av det stora hjärtat och förbättrar cirkulationen i dessa kroppsdelar.
Referenser
- R. F. Chapman: The insects, structure and function (4th ed, Cambridge University Press, 1998).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- B. Wigglesworth: Principles of insect physiology (7th ed, Chapman and Hall, 1972).
De bäst anpassade landdjuren tillhör de stora djurstammarna leddjur och ryggradsdjur. Nästan alla landryggradsdjur transporterar syre och koldioxid med hjälp av två olika system, lungor och blodomlopp. De flesta landlevande leddjur använder ett enda system för gastransporten, trakésystemet. Det är ett luftfyllt förgrenat rörsystem i vilket syre och koldioxid tranporteras från kroppsytan direkt till kroppens celler.
Bland leddjuren andas i stort sett alla insekter med trakéeer. Trakéer finns också hos mångfotingarna och hos många spindeldjur. Inom alla tre grupperna kan mycket små djur sakna trakéer och andas då bara genom huden. Trakésystem har kanske utvecklats tre gånger oberoende av varandra bland leddjuren. Trakésystemet transporterar syre och koldioxid hela vägen mellan luften och vävnaderna. Insekter kan därför klara sig med ett relativt enkelt uppbyggt blodomlopp, även om de har en hög syreförbrukning.

Hur fungerar trakésystemet?
Insekternas trakésystem är ett luftfyllt rörsystem som förgrenar sig i hela kroppen och transporterar syre och koldioxid mellan den omgivande luften och djurens celler. Rören är förstärkta med ringformade eller spiralformade lister med kitin. Kitin är en viktig komponent i insekternas yttre skelett. Trakéerna är inbuktningar från huden. Öppningarna på kroppsytan kallas spirakler. Det finns oftast ett par spirakler på vardera sidan av kroppen i varje segment, men antalet kan vara reducerat. I förrummet innanför spiraklerna finns ofta hår av kitin, motsvarande våra näshår, som håller förorenande partiklar borta. Där finns i regel också klaffar som periodvis kan stängas för att minska förlusterna av vatten i form av vattenånga. Detta är av särskilt stor betydelse för puppor, som ju inte kan dricka.
Luftsäckar är ofta kopplade till trakésystemet. Luftsäckar och grova trakéstammar kan tryckas ihop och utvidgas, när luften i de grövre delarna av trakésystemet byts ut vid ventilation, till exempel genom att bakkroppen omväxlande dras ihop och spänns ut i pumpande rörelser. På detta sätt transporteras ny syrerik luft in i trakésystemet. Transporten i de finare trakéerna sker dock genom diffusion. Många små insekter behöver förmodligen inte ventilera systemet alls. Hos dem sker transporten av gaserna bara med diffusion. Gasdiffusion innebär att molekyler av en viss gas, till exempel syre, rör sig från ett område med hög halt av denna gas (egentligen partialtryck) till ett område med lägre.
Trakeoler fungerar som "luftkapillärer"
De finaste trakégrenarna, motsvarande våra kapillärer, kallas trakeoler och ligger mycket nära cellernas membraner. Till skillnad från kapillärer slutar de blint. Avståndet mellan trakeolluften och cellernas syreförbrukande mitokondrier kan vara mindre än en tusendels millimeter. Videon överst på sidan visar hur komplicerat trakésystemet kan vara uppbyggt med både längsgående och tvärgående stammar och ett otal mindre grenar.

Hur andas vattenlevande insekter?
Det övervägande flertalet insekter drunknar förr eller senare i vatten. Detta gäller ägg, larver, puppor och vuxna. Trakésystemet hos dem fungerar inte i vatten. Det finns dock en hel del insekter som lever i sötvatten. De har alla kvar sitt gasfyllda trakésystem. Dessa insekter löser andningsproblemet på olika sätt.
En del vattenlevande insekter simmar helt enkelt upp till ytan för att utbyta syre och koldioxid med luften ovanför. Deras trakésystem har i regel bara ett par öppningar, spirakler, oftast belägna i bakänden. Ett exempel är larver av dykarskalbaggar och stickmyggor. De senare kan emellertid också ha så kallade trakégälar.
Hoppressbara gasgälar
Vissa vattenlevande insekter, till exempel ryggsimmare (släktet Notonecta) och vuxna dykarskalbaggar (släktet Dytiscus), har så kallade hoppressbara gasgälar. De tar, när de dyker, med sig en luftbubbla som försörjer dem med syre. Hos ryggsimmaren finns luften i rännor på buksidan, hos dykarskalbaggen under täckvingarna. Syre transporteras med diffusion från luftbubblan in genom trakésystemets öppningar, spiraklerna. När syrehalten sjunker i bubblan, transporteras löst syre från vattnet in i den med diffusion, vilket gör att den kan användas längre tid. Men samtidigt stiger bubblans kvävehalt. Detta gör att kväve diffunderar ut ur bubblan, vilket gör att den krymper och till slut försvinner. Då simmar insekten upp till ytan och hämtar en ny bubbla. Det finns en spindel, vattenspindeln (Argyroneta aquatica), som använder en hoppressbar gasgäle. Den har till och med ett luftförråd nere i vattnet som den kan besöka för att få syre. Men den måste fylla på förrådet då och då.

Icke hoppressbara gasgälar
Andra vattenlevande insekter har icke hoppressbara gasgälar, så kallade plastroner. De fungerar på samma sätt som hoppressbara gasgälar, med den skillnaden att luftutrymmet finns under en tät matta av vattenskyende hår. Vattnets ytspänning hindrar då vattnet från att tränga in i luftutrymmet, vilket gör att luftvolymen hela tiden är konstant. Eftersom volymen är konstant, kommer syrehalten att sjunka inne i plastronet, när insekten förbrukar syre. Då drivs nytt syre från vattnet in i plastronet med hjälp av diffusion och sedan vidare till trakésystemet. Men eftersom bubblans volym är konstant, förblir kvävehalten i bubblan konstant. Därför stannar kvävet kvar i bubblan. Insekter med plastron kan stanna mycket länge under vattnet, utan att besöka ytan, en del av dem i princip hur länge som helst. Ett exempel är vattenfisen (släktet Aphelocheirus), en liten skinnbagge. Vissa insektsägg är försedda med plastron i det yttre äggskalet. Hit hör spyflugornas ägg, som kan hamna i vätska när de läggs på ruttnande döda djur.
Slutna trakésystem och trakégälar
Alla de hittills nämnda vatteninsekterna har öppna trakésystem med spirakler. De finns emellertid andra vatteninsekter som har förlorat spiraklerna och därmed har slutna gasfyllda trakésystem. Små larver kan ta upp syre genom kroppsytan till ytligt belägna trakéer. Ett exempel är larver av knott (familjen Simuliidae) och av fjädermyggor (familjen Chironomidae). De senare har dessutom syrebindande hemoglobin, något som är mycket ovanligt hos insekter.
Större vattenlevande insekter med slutet trakésystem måste förstora sin kroppsyta på något sätt, annars kan de inte ta upp tillräckligt med syre. De ökar sin yta genom att ha så kallade trakégälar, bladformade eller trådformade utskott från kroppsytan eller baktarmen. Utskotten är försedda med ett tätt nätverk av trakéer. Syre transporteras genom ett tunt lager av vävnad till dessa trakéer. Exempel är larver av trollsländor (ordningen Odonata), nattsländor (ordningen Trichoptera) och bäcksländor (ordningen Plecoptera).
Fördjupning
- Läs artikeln Har myror ett hjärta? Om insekternas öppna cirkulation och människans slutna blodomlopp här ovanför.
- Du hittar mer om diffusion i nästa artikel Hur sker transporter i djurens kroppar? Diffusion och massflöde
Referenser
- R. F. Chapman: The insects, structure and function (4th ed, Cambridge University Press, 1998).
- D. R. Davis och J.-F. Landry: A review of the North American genus Epimartyria (Lepidoptera, Micropterigidae) with a discussion of the larval plastron (ZooKeys 183:37-83, 2012).
- R. W. Hill, G. A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- D. Iwan, M. J. Kamiński, och M. Raś: The last breath: a µCT-based method for investigating the tracheal system in Hexapoda (Arthropod Structure & Development 44:218-227, 2015).
- B. Wigglesworth: Principles of insect physiology (7th ed, Chapman and Hall, 1972).

Syre och koldioxid kan transporteras på två sätt: genom diffusion eller genom massflöde. Detsamma gäller alla andra ämnen som ska transporteras i kroppen. Kortväga transport, kortare än cirka 1 mm i vatten, kan ske med diffusion, medan långväga transport måste ske med massflöde. Detta kan du se när du blandar saft. Många ämnen, dock ej syre och koldioxid, kan också transporteras genom cellmembran med hjälp av proteiner, ofta energikrävande sådana.
Vid diffusion rör sig enskilda molekyler. Alla molekyler i vätskor och gaser är i ständig rörelse. Molekylerna rör sig oregelbundet och "studsar" ständigt mot varandra. Vid diffusion transporteras molekyler av ett visst slag (till exempel syremolekyler) genom molekylrörelserna i en viss riktning. Transporten drivs av en koncentrationsskillnad för just denna molekyl (i vårt fall syre). Syremolekyler lösta i vatten transporteras med diffusion från ett område med tätt mellan syremolekylerna (hög koncentration) till ett med glesare mellan syremolekylerna (låg koncentration). Notera att detta resonemang är något förenklat, när det gäller lösta gaser. (Det gäller dock strikt för oladdade molekyler av fasta ämnen, till exempel glukos och urea.)
Vid massflöde drivs en vätska med alla molekyler den innehåller från ett ställe till ett annat. Massflödet drivs nästan alltid av en tryckskillnad. Vätskan flödar från ett område med högt tryck till ett område med lågt. Vattnet som kommer ur vattenkranen rör sig med hjälp av massflöde. Blodet i cirkulationssystemet, urinen i njurarna, innehållet i magtarmkanalen och luften i lungorna rör sig också med hjälp av massflöde. Dessa långväga transporter kan bara ske med massflöde och i princip hur långt som helst: från vattentornet till vattenkranen eller från hjärtat till bakänden hos en blåval.
Om du ska blanda saft och vatten kan du försiktigt skikta vattnet ovanför saften, så att du får ett saftlager underst och ett vattenlager överst. Sedan kan du vänta på att saftmolekyler ska röra sig från saften (där koncentrationen av saftmolekyler är hög) till vattnet (där koncentrationen är 0, där det inte finns några saftmolekyler). Rätt snart börjar gränslinjen mellan saft och vatten bli otydlig. Det beror på att saftmolekylerna rör sig med diffusion in i vattenskiktet. Men du får vänta väldigt länge, innan saften blandats färdig på detta sätt. Den vane saftblandaren rör naturligtvis om i glaset och åstadkommer då tryckskillnader som på några ögonblick blandar saften med vattnet. Den vane saftblandaren använder sig av massflöde.
Fördjupning om diffusion
Tänk dig två likadana vattenfyllda behållare som är åtskiljda av ett plant membran, en barriär. I den vänstra har vi en lösning med hög koncentration av ämnet X, i den högra en lösning med låg. Ämnet X består av oladdade molekyler av ett fast ämne lösta i vatten. För joner, som har elektrisk laddning, och för gasmolekyler bli situationen mer komplicerad. Koncentrationen av X definieras som antalet molekyler per volymenhet vätska och mäts med enheten mol/liter (gram/liter hade också gått bra). Membranet släpper igenom X, men är ogenomsläppligt för vatten.

Genom de så kallad termiska molekylärörelserna rör sig X-molekylerna ständigt slumpmässigt och krockar ofta med varandra. Då och då leder detta till att en X-molekyl passerar membranet. Eftersom X-koncentrationen är högre i den vänstra behållaren än i den högra, kommer fler molekyler passera membranet från vänster till höger, än från höger till vänster. Vi får således en nettotransport från vänster till höger. Denna nettotransport är diffusionen och mäts i antal molekyler per tidsenhet eller mol/sekund. Diffusionen pågår tills dess koncentrationerna på båda sidor av membranet blir lika stora. Vi har då nått diffusionsjämvikt.
Intuitivt kan vi kanske föreställa sig att följande gäller:
- Diffusionen (M mol/sekund) ökar, om skillnaden i koncentration mellan den vänstra behållaren (Cvä mol/liter) och den högra (Chö mol/liter) ökar. Då kommer nämligen skillnaden mellan antalet X-molekyler, som går åt höger, och antalet, som går åt vänster, bli större.
- Diffusionen ökar om membranets yta (A cm2) ökar, eftersom detta ger fler möjligheter för X-molekyler att passera membranet.
- Diffusionen minskar om membranets tjocklek (L cm) ökar, eftersom det då blir svårare för X-molekylerna att passera barriären.
- Diffusionen minskar om X har en högre molekylvikt, eftersom stora molekyler rör sig långsammare än små.
- Difffusionen minskar om X-molekylerna har svårt att lösa sig i membranet. Ett cellmembran är ett exempel. Det består huvudsakligen av fettämnen (fosfolipider). Vattenlösliga molekyler stöts bort av fettämnena och löser sig mycket dåligt i membranet. Därför blir deras diffusion genom membranet mycket liten.
- Diffusionen minskar om lösningarnas temperatur (K) minskar, eftersom X-molekylerna då rör sig långsammare. Temperaturen har dock en relativt liten effekt på diffusionen.
För den matematiskt intresserade kan allt detta sammanfattas som:

D är diffusionskoefficienten. Dess storlek minskar, om molekylvikten för X ökar, om X har svårare att lösa sig i membranet och om temperaturen minskar.
I lungorna är förhållandena mer komplicerade, men de allmänna principerna ovan gäller. Ett problem är att alveolernas (lungblåsornas) inre yta är buktig, inte plan. Det matematiska uttrycket ovan blir då mer komplicerat. Men detta brukar vi bortse ifrån inom fysiologin. Två andra problem är dels att syre och koldioxid är gaser, dels att diffusionen i lungorna sker mellan en gas, alveolluften, och en vätska, lungvävnaden. Dessa problem kan vi inte bortse ifrån. De leder till att formeln ovan måste modifieras. Men jag diskuterar dem inte här. Ytterligare ett problem är att diffusionbarriären i lungorna är komplicerat uppbyggd och består av ett vätskeskikt, alveolceller med cellmembran, vävnadsvätska, kapillärväggens celler med cellmembran, blodplasman och de röda blodkropparnas cellmembran.
En stor sfär har mycket mindre yta än många små sfärer med samma totala volym. Om våra lungor hade varit enkla säckar, som grodornas, hade de haft en mycket liten diffusionsyta per volym lunga. I och med att lungorna innehåller ett otal mycket små alveoler, genom vilkas väggar diffusion kan ske, blir den totala diffusionsytan per volym lunga enormt mycket större.
Fördjupning
Artikeln Kan människor andas vatten? Om våra luftvägar och lungor finns här ovanför.

Sälar är däggdjur som andas med lungor precis som vi människor. Det betyder att de måste hålla andan, när de dyker efter föda. Sälarna kan dyka flera hundra meter under ytan och en dykning kan vara flera minuter!
Människan, som också är ett däggdjur, kan inte jämföra sig med sälar. Många av er kan förmodligen simma. Ni har säkert vid flera tillfällen tävlat med era kompisar i dykning. Själv är jag uppvuxen i Turkiet och har lekt mycket i det djupa och underbara Medelhavet. Jag har aldrig varit någon höjdare på dykning och brukade komma sist på tävlingarna. Det bästa jag och mina kompisar kunde prestera var några ynkliga meter, kanske mellan tre och fyra. Men när vi pratade om det, speciellt framför tjejer, så brukade det vara 9-10 meter. Det finns människor som kan fridyka ner till 60 meters djup eller mer, men även detta är inget jämfört med sälarnas dykförmåga. Weddellsälen är känd för att kunna dyka ända ner till 600 meters djup och för att kunna stanna under vatten i mer än 1 timme.
Sälarna är trots allt anpassade till ett liv i vatten, medan vi är landanpassade. Du skulle kunna tro att sälarnas lungor är mycket större än våra, men det är inte sant. Det skiljer faktiskt inte så mycket mellan en sällunga och människolunga, om du tar hänsyn till kroppsvikten. En viktig skillnad är att sälarnas bröstkorg är mer elastisk än människans. Detta är viktigt, eftersom trycket ökar enormt med djupet. Trycket på 100 meters djup cirka 10 gånger större än trycket vid havsytan. Vid detta tryck kan sälens bröstkorg och lungor pressas ihop utan några problem. Sälarnas lungor används inte som syrgasförråd vid dykning, tvärtom är det bra att de pressas ihop. Då ger de nämligen mindre flytkraft, vilket naturligtvis är bra när de dyker nedåt. Dessutom blir risken mindre att för mycket kväve löses i kroppsvätskorna på grund av det höga trycket. I den situationen skulle kvävet kunna bilda gasbubblor i vävnaderna, när sälen återvänder till ytan och trycket minskar. Detta kallas dykarsjuka. Mänskliga dykare som andas från en gastub måste stiga upp till ytan långsamt för att undvika den farliga dykarsjukan.
Eftersom syrgas inte kan tas upp från omgivningen, så måste dykande djur använda sina syreförråd. En viktig sådan depå är myoglobinet som finns i skelettmusklerna (de viljestyrda musklerna) och ger dem deras röda färg. Myoglobin är ett protein besläktat med blodets röda hemoglobin och har, liksom hemoglobinet, förmåga att binda syremolekyler. Myoglobinet är särskilt viktigt när syrehalten blir låg i blodet. Sälar har en oerhört stor mängd myoglobin i musklerna. Sälmuskler är mycket mörkt röda av myoglobin. Myoglobinet används vid dykning som syreförråd av skelettmusklerna.
Sälarnas blod innehåller också ett större syreförråd än människans blod. Detta gäller särskilt de sälar som kan stanna mycket länge under vattnet. En ökad syremängd i blodet kan åstadkommas genom att blodet har en högre halt av röda blodkroppar och därmed en högre halt av syre bundet till hemoglobin. Ett stort syreförråd kan också åstadkommas genom att blodvolymen, och därmed den totala hemoglobinmängden i kroppen, är stor i förhållande till kroppsvikten. Många dykande sälar förvarar dessutom syrerika röda blodkroppar i mjälten, medan de andas luft. När de dyker ökar de blodets syreförråd genom att överföra dessa blodkroppar till blodet.
En annan mycket viktig egenskap är att sälar kan sänka sin hjärtslagsfrekvens (pulsen) och samtidigt strypa blodflödet till stora delar av kroppen. Detta gör de nästan bara vid mycket långa dyk. Viktiga organ som hjärtat och hjärnan får då det blodflöde de behöver, medan andra organ, till exempel mag-tarmkanalen och skelettmusklerna, får en mycket litet blodflöde. Eftersom de senare organen kan utvinna energi anaerobt utan syre under bildning av mjölksyra, så behöver de inte så mycket syre på kort sikt. På så sätt används syreresurserna på ett sparsamt sätt. Den sänkta hjärtslagsfrekvensen förhindrar att strypningen av blodflödet leder till en farlig höjning av blodtrycket. Den låga pulsen gör dessutom att hjärtats arbetsbelastning minskar och därmed dess syrebehov.
Referenser
R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).

Den som myntade uttrycket "som en fisk på torra land", såg nog inte lungfiskarna framför sig. Dessa exceptionella fiskar har en förmåga att överleva upp till ett år utan tillgång till vatten. Vanliga fiskar, som bara andas med gälar, skulle kvävas om de blev berövade sin naturliga miljö, vattnet, medan lungfiskarna till och med har utvecklat en förmåga att andas med lungor.
Forskare tror att dessa fiskar utvecklat lungor, eftersom de har levat i väldigt växlande miljöer, i vilka vattenmassorna lätt kunde torka ut och exponera sina invånare för omgivande luft. Om någon art kunde utveckla lungor att användas vid luftandning, skulle det utgöra en stor fördel i en sådan miljö. Förutom sina lungor, äger lungfiskarna ytterligare en anpassning till växlande miljöer. De har speciella skaftade fenor som kan ha gjort det möjligt för en del lungfiskar att röra sig till en ny vattensamling, när den de levt i torkar ut. Fenorna är dock starkt reducerade hos två av de nutida släktena.
Numera representeras lungfiskarna av tre släkten som alla återfinns i instabila sötvatten. Dessa tre släkten förekommer på tre olika kontinenter: Afrika, Sydamerika och Australien. Den australiska lungfisken (Neoceratodus forsteri) måste alltid andas vatten med gälarna, men kan skaffa sig extra syre genom att andas luft med lungorna, om syrehalten sjunker i vattnet. De sydamerikanska (släktet Lepidosiren) och afrikanska (släktet Protopterus) lungfiskarna dör om de inte får tillfälle att andas luft.

Både de afrikanska och de sydamerikanska lungfiskarna kan gå in i ett vilstadium, om deras vattenmiljö torkar ut helt. De gräver då ner sig i sjöbottnen. De afrikanska lungfiskarna utsöndrar dessutom ett slags slem som hårdnar och bildar en kokong runt fisken. Den enda kontakt med yttervärlden, som då kvarstår, är ett litet andningshål i kokongen vid huvudet, genom vilket fisken får luft till sina lungor. Kokongen skyddar fisken mot vattenförluster. Eftersom fisken varken dricker eller äter, är det av största vikt att vattenbalansen bibehålls.
När näringstillförseln upphör krävs det också att lungfisken hushållar med sina energireserver. Den minskar sin ämnesomsättning med 90 procent, vilket gör att dess energiförråd i form av fett och muskelproteiner räcker längre. Genom denna minskning av ämnesomsättningen minskar också vattenförlusterna. Eftersom det är ämnesomsättningen som kräver syrgas, behövs inte längre samma mängd syrgas. Då kan lungandningen minska, något som minskar vattenförlusterna via andningsluften.
Ytterligare en anpassning, som gjort det möjligt för lungfiskarna att gå in i ett vilstadium, är deras slutprodukt vid proteinnedbrytningen. De flesta vattenlevande djur bildar ammoniak som slutprodukt, men dessa djur måste göra sig av med denna lika fort som den bildas, eftersom ammoniak är mycket giftigt. De flesta fiskar gör sig lätt av med den vattenlösliga ammoniaken via gälarna. Så gör lungfiskarna också så länge de befinner sig i vatten Eftersom gälar inte fungerar i luft måste lungfisken lösa slutproduktsproblemet på annat vis, när den bara andas luft. Den gör det genom att i stället för ammoniak bilda urea (urinämne) som slutprodukt. Urea är för övrigt även slutprodukt hos oss däggdjur och inte alls lika giftig som ammoniak. Lungfisken kan låta urea ansamlas i kroppen tills dess koncentration når otroliga värden, utan att bli förgiftad.
Alla dessa anpassningar till landliv har gjort det möjligt för en fisk att överleva upp till ett år, kanske längre, på torra land.
Referenser
- Q. Bone och R.H. Moore: Biology of fishes (3rd ed, Taylor & Francis, 2008).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K.V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).

Hur gör fåglar egentligen för att andas när de ligger inneslutna i ägg med hårt skal? Ett fågelägg innehåller nästan allting som ett embryo behöver för att utvecklas till en färdig fågelunge. Det enda som inte får plats i ägget är syrgas. Men äggskalet är fullt av mikroskopiskt små porer genom vilka gaser av olika slag kan passera in och ut ur ägget.
Genom porerna tar kycklingfostret upp det syre som den behöver. Genom porerna kan fostret också göra sig av med den koldioxid som bildas som en avfallsprodukt. Gaserna transporteras med så kallad diffusion. Diffusion drivs normalt av koncentrationsskillnader, för gaser av så kallade partialtryckskillnader. Eftersom fostret förbrukar syre blir syrekoncentrationen, och därmed partialtrycket för syre, lägre inuti ägget än utanför och syremolekyler diffunderar därför in i ägget. Ett vanligt hönsägg har ungefär 10 000 porer! Varje por har en diameter på 0,017 millimeter och den totala porytan på ett ägg är 2,3 kvadratmillimeter.

Hönsäggets uppbyggnad
Innerst finns gulan. Det är den som är äggcellen och den är omgiven av det mycket tunna vitellinmembranet. Större delen av gulan är fylld av upplagrad näring åt embryot i form av tätt packade gulekorn. Groddskivan syns som en vit fläck på gulans yta. I den finns äggets kärna och det är den som utvecklas till ett embryo i ett befruktat ägg. I övrigt består gulan av flera lager av omväxlande vitaktig gula (klart gul på bilden) och gulare gula (mörkare på bilden). Den vitaktiga gulan bildar en pelare under groddskivan.
Utanför gulan finns vitan som även den utgör ett förråd av näring och vatten åt embryot. Vitan fungerar också som en kudde som skyddar det tidiga embryot från stötar. Dessutom har vitan bakteriedödande egenskaper. Den innehåller ett enzym, lysozym, som bryter ner cellväggarna hos många bakterier. Vitan består av flera lager med omväxlande trögflytande och mera lättflytande vita. Den trögflytande vitan närmast gulan bildar de båda äggsnoddarna (mörkt gråa) som fungerar som fjädrande upphängningsanordningar för gulan.
Utanför vitan finns de båda tunna skalmembranerna som ligger tätt ihop under skalet, utom i äggets trubbiga ände. Här finns en luftkammare mellan de båda skalmembranerna. Ytterst ligger skalet (brunt på bilden) som till stor del består av kalciumkarbonat. Skalet är perforerat av en mängd små porer. Gulan har lägst densitet ("täthet") i området kring groddskivan och högre vid den motsatta polen. Om du vänder ett ägg som ligger på sidan kommer gulan därför att långsamt vrida sig tills groddskivan åter ligger överst.
Kycklingens andning
Ett kycklingfoster kan förstås inte andas genom lungorna. Det har ju inga fungerande lungor ännu. I stället tar det upp syret med hjälp av en av fosterhinnorna. Inuti ägget ligger kycklingfostret omgiven av flera fosterhinnor. En av dessa hinnor kallas allantois. Allantois växer ut som en utbuktning från fostrets tarm. Först är allantois en ihålig blåsa, sedan ligger den som en tillplattad ballong längs skalets insida, där den växer ihop med en utanför den liggande fosterhinna som kallas korion. Det är här gasutbytet sker. Blod från kycklinfostret cirkulerar ut i allantois, tar upp syre och avger koldioxid. Det syrerika blodet strömmar sedan tillbaka in till fostret.
Under den sista utvecklingstiden händer det mycket i ägget. Ungefär 28 timmar innan kläckningen pickar kycklingen hål på äggets luftkammare, som nu ökat i volym till cirka 10 milliliter. Nu börjar lungorna långsamt att fungera och kycklingen kan andas av den luft som finns i luftkammaren. Under en övergångsperiod får den syre både via lungorna och via allantois. Sedan börjar kycklingen picka hål på äggskalet. Medan den kämpar för att ta sig ut ur skalet, övergår den till att andas endast med hjälp av lungorna.
Ett hönsägg ruvas i ungefär 21 dagar. Under den här tiden tar ägget upp syre, samtidigt som det avger koldioxid och vattenånga till omgivningen. Det är ganska stora mängder gas det handlar om. Sammanlagt tar ägget upp 6 liter syre och avger 4,5 liter koldioxid. Eftersom skalets porer släpper igenom alla slags gaser, förlorar ägget också en hel del vatten i form av vattenånga, totalt 11 liter vattenånga. Vid äggets trubbiga ände finns en ficka med luft. Det är denna luftkammare som syns som en fördjupning på ett kokt ägg. Luftkammaren ligger mellan de båda skalmembranerna och ökar i storlek allteftersom vatten avdunstar från ägget. De båda skalmembranerna finns under skalet runt hela ägget och syns när du skalar frukostägget, men de ligger tätt ihop utanför luftkammaren.
Referenser
- R.L. Dorit, W.F. Walker Jr och R.D. Barnes: Zoology (Saunders, 1991).
- H.W. Manner: Elements of comparative vertebrate embryology (Macmillan, 1964).
- A. Reizis1, I. Hammel2 och A. Ar: Regional and developmental variations of blood vessel morphometry in the chick embryo chorioallantoic membrane (Journal of Experimental Biology 208:2483-2488. 2005).
- K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).
- W. Shumway och F.B. Adamstone: Introduction to vertebrate embryology (5:e upplagan, Wiley, New York, 1962).
Njurar och utsöndring – Om salt- och vattenbalansen i kroppen

Våra njurar betraktas ofta bara som en slags biologiska sopnedkast för "slaggprodukter". Detta är mycket långt från sanningen. Njurarna har en mängd andra livsnödvändiga funktioner.
Människans njurar avger visserligen slutprodukter av ämnesomsättningen, bland annat urinämne och urinsyra. Men de har dessutom en hel rad andra funktioner, som gör att kroppsvätskornas sammansättning kan regleras med extrem noggrannhet. Njurarna reglerar mängderna och koncentrationerna i kroppen av bland annat natriumjoner och kloridjoner (alltså salt, mera exakt koksalt), vatten, kalciumjoner, kaliumjoner, fosfatjoner och ett flertal andra ämnen. De håller kroppsvätskornas pH på en nära nog konstant nivå genom att styra utsöndringen av vätejoner och vätekarbonatjoner. De omvandlar och utsöndrar dessutom en mängd kroppsfrämmande ämnen, till exempel gifter och läkemedel.

Nederst ses en närbild av en glomeruluskapillär från bilden överst på denna sida. Men du ser inte det cellskikt som avgränsar kapillären, det så kallade endotelet. Endotelet är helt täckt av celler tillhörande kapselns inre epitel. Dessa celler är trädlikt förgrenade med ett otal fingerlika utskott. Kapselns inre epitel är således svept runt varenda kapillär i hela glomerulus. Mellan de fingerlika utskotten finns tunna hinnor av så kallade glykoproteiner (som inte syns på bilden). Dessa hinnor filtrerar bort blodets stora molekyler, plasmaproteinerna, från primärurinen. Blodets celler, blodkropparna, filtreras bort av kapillärernas endotel. Primärurinen består således bara av små molekyler, joner och vatten. Skalstrecket motsvarar 5 µm (0,005 mm).
Filtrering, reabsorption och sekretion
I människans njurar och de flesta andra djurs utsöndringsorgan bildas den så kallade primärurinen genom att vätska filtreras genom ett membran. Filtret släpper inte igenom celler och inte mycket stora molekyler, som proteiner. Det är alltså samma princip som när du lagar kaffe, då kaffekornen inte passera filtret. Filtrering av primärurin drivs dock inte av trycket skapat av tyngdkraften, som i en vanlig kaffebryggare, utan av ett annat tryck. Det är ofta blodtrycket, som pressar vätska från blodet genom filtret, varvid celler och stora molekyler inte passerar. Filtreringen fungerar alltså som i sådana små kaffebryggare, i vilka du med handkraft pressar vatten genom kaffet och ett filter.

Från kapseln drivs primärurinen genom ett rörsystem, tubulus. Tubulus består av flera avsnitt med olika funktioner. I dessa avsnitt modifieras primärurinen på olika sätt. Blodet lämnar glomeruluskapillärerna i den bortförande arteriolen. Eftersom blodet fortfarande har en hög syrehalt är detta kärl en arteriol, inte en venol. Det för blodet till nya kapillärer, dels i njurens yttre del, barken, dels i dess inre del, märgen. Dessa kapillärsystem utbyter vatten och lösta ämnen med tubulus.
Reabsorption är transport från tubulus till kapillärerna. Sekretion är transport fråm kapillärerna till tubulus. Noterbart är att cirka två tredjedelar av av den flltrerade primärurinen (cirka 180 l/min) reabsorberas redan i tubulus första avsnitt, proximala tubulus. Noterbart är också att njurarna har två kapillärsystem kopplade i serie, utan att blodet emellan dem passerar hjärtat. Blodet från det andra kapillärsystemet förs via venoler till vener. Den färdiga urinen töms i ett hålrum, njurbäckenet, på njurens konkava sida. Urinen modifieras inte vidare när den därifrån lämnar kroppen via de båda njurarnas urinledare, urinblåsan och urinröret.
Glomerulus och Bowmans kapsel utgör tillsammans en njurkorpuskel. En njurkorpuskel tillsammans med sin tubulus utgör ett nefron. Grovt räknat finns det 1–2 miljoner nefron i de båda njurarna.
Primärurinen rinner sedan vidare genom en utförsgång, ofta kallad tubulus. I de flesta, kanske alla, utsöndringsorgan modifieras primärurinen, när den passerar genom utförsgången. Detta sker genom att lösta ämnen och vatten transporteras genom gångens vägg, något som kan ske på två sätt. Vid reabsorption återvinnes molekyler och joner från primärurin till kroppsvätska. Vid sekretion förs molekyler och joner från kroppsvätska till primärurin.
Hos enkel byggda djur mynnar utförsgången på kroppsytan. Ofta är den dessförinnan vidgad till en urinblåsa som lagrar urin. Hos människan och andra ryggradsdjur passerar urinen genom ett mer komplicerat gångsystem innan den lämnar kroppen.

Innanför märgen finns ett hålrum, njurbäckenet (pelvis) som samlar upp den färdiga urinen. Genom vandrande sammandragningar av glatt muskulatur (peristaltik) i urinledaren (urether) pressas urinen vidare till urinblåsan. Bäda njuranas urinledare mynnar i urinblåsan. När urinblåsans glatta muskulatur kontraheras pressas urin in i urinröret (urethra). När urinrörets båda slutmuskler relaxeras lämnar urinen kroppen. Den ena slutmuskeln består av icke viljestyrd glatt muskulatur, den andra av viljestyrd skelettmuskulatur.
Njuren försörjs med blod via njurartären och blodet lämnar den via njurvenen (båda kärlen är avskurna på bilden). Fantastiskt nog mottar de båda njurarna cirka 20 procent av det blod som pumpas ut av hjärtat i det stora kretsloppet, ungefär 1 liter blod/minut i vila. Detta är en av flera viktiga faktorer, som gör att njurarna så precist kan reglera kroppsvätskornas sammansättning.
Passiv reabsorption och sekretion i tubulus
Reabsorption och sekretion kan ske passivt, utan energitillförsel, genom diffusion eller osmos.
Diffusion innebär att molekyler eller joner av ett visst ämne spontant drivs från ett område med hög koncentration av just detta ämne till ett med lågt. Joner, som har en elektrisk laddning, kan även spontant drivas av en elektrisk spänning.
Osmos innebär att vatten spontant tenderar att röra sig från ett område med låg totalkoncentration av lösta ämnen till ett område med högre, förenklat uttryckt från ett område med hög "vattenkoncentration" till ett område med låg. Osmolariteten (mOsmol per liter) är ett mått på en lösnings benägenhet att dra till sig vatten med osmos.
Joner utövar en mindre osmotisk effekt än oladdade molekyler. Enkelt uttryckt beror detta på att positivt laddade joner har en tendens att binda till negativa joner i en lösning. En saltlösning har därför en lägre osmolaritet än summan av dess jonkoncentrationer. En urinämneslösning har däremot samma värde på koncentration och osmolaritet, åtminstone i låga koncentrationer.
Passiv transport kan ske genom proteinkanaler i de cellmembraner, som omger cellerna i utförsgångens vägg, rakt igenom dessa cellers cellmembran (endast fettlösliga ämnen) eller mellan dessa celler. Molekyler och joner som tranporteras passivt kan jämföras men stenar, som rullar ner för en backe.
Aktiv reabsorption och sekretion i tubulus
Reabsorption och sekretion kan också ske aktivt under tillförsel av energi. Primär aktiv transport av oorganiska joner kan åstadkommas med hjälp av membranproteiner, som drivs av den energirika molekylen ATP. Jonen rör sig då i motsatt riktning mot den riktning, i vilken den drivs av diffusionen, något som kräver tillförsel av energi. Jonen kan då jämföras med en sten, som rullas uppför en backe.
ATP-driven aktiv transport av en jon kan sedan indirekt driva den sekundära aktiva transporten av andra ämnen. Dessa ämnen kan vara organiska molekyler, till exempel glukos (druvsocker), eller oorganiska joner, till exempel kalcium. Då fungerar koncentrationsskillnaden över utförsgångens vägg för en jon, i regel natriumjonen, som energikälla. Ett protein i ett cellmembran transporterar både natriumjonen och en annan molekyl eller jon. Natriumjonen transporteras i sin diffusionsriktning, vilket frigör energi. Molekylen eller jonen transporteras mot sin diffusionsriktning, vilket kräver energi.
Överskottett av natriumjoner på membranets ena sida är ett energiförråd, precis som vattnet i en kvarndamm. Natriumtransporten fungerar som vattnet i ett vattenfall. Membranproteinet fungerar som ett kvarnhjul i en vattenkvarn. Natriumflödet driver proteinet, precis som vattenfallet driver hjulet. Hjulet driver ett tranportband, som transporterar stenar uppför en backe. Stenarna motsvarar den andra molekylen eller jonen. På detta sätt reabsorberas normalt all glukos från primärurinen.
Styrning av kroppsvätskornas sammansättning
Bildning av primärurin genom filtrering förekommer hos många djurgrupper, till exempel rundmaskar, många mollusker, många kräftdjur samt nästan alla ryggradsdjur. Forskarna vet dock mycket lite om exkretionsorganen hos de flesta djurgrupper. Ofta filtreras stora mängder primärurin som sedan reabsorberas till allra största delen. Detta är naturligtvis energimässigt kostsamt och vi kan fråga sig vad det medför för fördelar.
Med en hög filtreringstakt har vi en icke selektiv mekanism genom vilken alla lösta ämnen kan tillföras urinen, utom proteiner och andra mycket stora molekyler. Med en specifikt reglerbar och hög reabsorptionstakt har vi en selektiv mekanism genom vilken utsöndringen av varje enskilt ämne mycket snabbt kan ökas eller minskas. Dessutom sker aktiv sekretion av vissa ämnen, till exempel gifter, som behöver avlägsnas extra snabbt ur kroppen. Därmed får vi en mycket effektiv reglering av kroppsvätskornas sammansättning.
Människans njurar fungerar på ovan beskrivna sätt. Filtreringen (den så kallade GFR) är cirka 180 liter primärurin per dygn (!) hos en person på 70 kg. Cirka 99 procentav denna volym reabsorberas, mer vid vattenbrist och mindre vid vattenöverskott i kroppen. Vi hade avlidit av vattenbrist inom några timmar, om vi inte reabsorberade den överväldigande merparten av primärurinen.
Reabsorption och sekretion hos människan
Våra njurar är, som nämnts ovan, inte biologiska sopnedkast. De källsorterar i stället de ämnen som finns i kroppsvätskorna. Därvid avlägsnar de med urinen mer av de ämnen, som riskerar att nå alltför höga halter i kroppen, och mindre av de ämnen, som riskera att nå alltför låga halter i kroppen. På så sätt regleras till exempel halterna och mängderna av vatten, natriumjoner och kloridjoner i kroppsvätskorna mycket effektivt.
Vissa nyttiga ämnen, som vi behöver vid ämnesomsättningen, reaborberar vi helt och hållet. De finns därför normalt inte i den slutliga urinen. Andra ämnen, som är slutprodukter vid ämnesomsättningen, avges i stora mängder med den slutliga urinen. Till exempel så återvinnes glukos (druvsocker) helt och hållet från primärurinen, medan stora mängder urinämne (urea) avges med slutliga urinen. Glukos är en energikälla, som bryts ner vid cellandningen under bildning av koldioxid, vatten och enenrgirikt ATP. ATP driver kroppens energikrävande processer. Urinämne är en slutprodukt vid nedbrytningen av proteinernas aminosyror. Det är giftigt i alltför höga halter. Vatten, salt och många andra ämnen utsöndras i stora mängder, när de finns i överkott i kroppen, och i små mängder, när det råder brist i kroppen.
Urinens osmolaritet. Osmoreglering
Urin hos djur kan vara isoosmotisk med samma osmolaritet som kroppsvätskorna. Den kan även vara hypoosmotisk med lägre osmolaritet än ("mindre koncentrerad än") kroppsvätskorna. Slutligen kan den vara hyperosmotisk med högre osmolaritet än ("mer koncentrerad än") kroppsvätskorna. Människan kan producera alla dessa tre typer av urin, vid vattenöverskott hypoosmotisk och vid vattenbrist hyperosmotisk. Osmoreglering är regleringen av kroppsvätskornas osmolaritet.
Havslevande ryggradslösa djur har vanligen samma osmolaritet i kroppsvätskorna som i havsvattenet. De producerar därför nästan alltid en isoosmotisk urin och undviker på så sätt osmotisk stress.

Den saltvattenslevande sillen (Clupea harengus), nederst, måste dricka stora mängder saltvatten. Annars hade den krympt och dess celler dött.
Det är omöjligt för djur att ha samma låga osmolaritet i kroppsvätskorna som i sötvatten. Sötvattensdjur har därför en högre inre osmolaritet än sötvattnet, vilket innebär att de ständigt tar upp vatten med osmos, särskilt i gälarna. För att inte svälla upp och spricka, måste de därför ständigt avge enorma mängder hypoosmotisk urin. För att upprätthålla sin inre osmolaritet måste de dessutom ta upp salt från vattnet med energikrävande transport, vanligen i gälarna. De har dock anpassat sig till sötvattnet genom att sänka sin inre osmolaritet, vilket minskar den osmotiska stressen.
Havslevande benfiskar (teleoster) har en avsevärt lägre osmolaritet i kroppsvätskorna än havsvattnet, kanske på grund av de härstammar från sötvattenslevande fiskar och av okänd anledning inte kunnat höja sin inre osmolaritet till havsvattnets nivå. Därför förlorar de ständigt vatten med osmos, särskilt i gälarna. De avger en mycket sparsam isoosmotisk urin. Men de måste kompensera för vattenförlusten genom att dricka havsvatten. Detta ger en stor saltbelastning. De måste därför göra sig av med saltet med energikrävande tranport i gälarna. Det stora flertalet nu levande fiskar är benfiskar, däribland nästan alla våra vanliga matfiskar.
Landlevande djur lider ofta brist på vatten, eftersom vatten avdunstar från kroppytan och avges som vattenånga via utandningsluften. De avger därför ofta en sparsam isoosmotisk urin. Naturligtvis hade det varit bättre för dem att avge en hyperosmotisk urin och på så sätt spara ännu mer av kroppens vatten. Men märkligt nog är det bara tre djurgrupper, som är så väl anpassade till landliv, att de kan avge en hyperosmotisk urin. Dessa grupper är insekter, däggdjur och fåglar.
Djur som saknar utsöndringsorgan
Många djur saknar helt urinproducerande utsöndringsorgan (exkretionsorgan). Dessa djur är ofta mycket små. De kan också vara stora djur med sina kroppsceller belägna huvudsakligen i anslutning till huden och mag-tarmkanalens vägg. Det senare gäller nässeldjur (som maneter och koraller), kammaneter och tagghudingar (som sjöstjärnor och sjöborrar). Hos dessa djur utan utsöndringsorgan är transportavstånden inuti dem vanligen korta. De kan därför avge sina utsöndringsprodukter antingen genom huden till omgivningen eller genom mag-tarmkanalens vägg till dess hålrum, ofta med diffusion eller osmos.
Stammen Xenacoelomorpha, en djurgrupp som anses var ursprungligast bland de tvåsidiga (bilateralsymmetriska) djuren, saknar utsöndringsorgan. Nästan alla andra tvåsidiga djur har utsöndringsorgan. Vissa små sådana djur har dock förlorat dessa organ. Som nämnts ovan saknas utsöndringsorgan också hos tagghudingarna, dock bara hos de vuxna djuren.
Protonefridier, metanefridier och njurar
Det finns två ursprungliga typer av utsöndringsorgan hos djur, protonefridier och metanefridier. Det är oklart om dessa exkretionsorgan uppkommit en eller flera gånger under djurens evolution. Ibland har utförsgången ett annat ursprung än nefridiet. Organet kallas då nefromixium. Protonefridier tros vara den ursprungligaste typen.

Protonefridier, är slutna rörformade inbuktningar från huden. I den slutna änden är de försedda med en eller flera celler med ett eller några gissel (flageller) eller med många flimmerhår (cilier). Via en utförsgång till hudens yta avger de urin till omgivningen. Protonefridiernas funktion är dåligt känd. Men forskare tror att gisslens och flimmerhårens rörelser åstadkommer ett undertryck, som suger in vätska i dem genom filtreringsmembraner runtom dem. Stora molekyler, som proteiner, passerar inte filtren. Protonefridier finns främst hos små djur, särskilt de som saknar kroppshåla eller bara har en litet så kallat coelom (sekundär kroppshåla). Exempel på sådan djur är plattmaskar, hjuldjur och några ringmaskar.

Metanefridier skiljer sig från protonefridierna framför allt genom att de börjar med en öppen tratt (nefrostomen). Tratten är oftast på insidan försedd med flimmerhår (cilier) som driver vätskan in i den. Tratten är i regel belägen i kroppshålan, coelomet. Om filtrering sker, kan förmodlgen coelomets vägg i en del fall fungera som filter. Tratten är förbunden med ett rör som mynnar på kroppsytan. Rörets är ofta uppdelat i olika delar med olika struktur och olika funktion. I vissa rördelar sker förmodligen reabsorption och sekretion av lösta ämnen och vatten. Men metanefridiernas funktion är dåligt utforskad.
Komplicerat byggda djur har ofta metanefridier som är starkt modifierade med mer eller mindre effektiv filtrering. Dessa organ kallas vanligen njurar.
Blötdjurens njurar
Njurarna hos nästan alla vuxna blötdjur är mer eller mindre starkt modifierade metanefridier. De börjar oftast i den del av coelomet som omger hjärtat. Ofta finns det kärl som för hemolymfa till njurarna. Hos blötdjur och andra djur med öppet kärlsystem fungerar hemolymfan som blodet gör hos djur med slutet kärlsystem. Njurarnas utförsgångar kan vara vidgade till en urinblåsa. De mynnar i den så kallade mantelhålan, från vilken den färdiga urinen lämnar djuret. Reabsorption och ibland även sekretion sker i utförsgångarna. En del mollusker, till exempel bläckfiskar, kan filtrera primärurin med högt tryck, något som gör utsöndringen effektivare.
Antennkörtlar och maxillkörtlar hos kräftdjur
Många kräftdjur har antennkörtlar eller maxillkörtlar, som kanske är omvandlade metanefridier hos vilka tratten slutits. En antennkörtel börjar med ändsäcken (coelomsäcken). In i den filtreras sannolikt proteinfri hemolymfa. Därefter följer den så kallade labyrinten, en rörformad nefridiekanal och till sist urinblåsan som mynnar på huvudet vid den andra antennen. Filtratet modifieras med reabsorption och sekretion på sin väg genom körteln. Notera att många kräftdjur således urinerar med huvudet.
Malpighiska kärl hos insekter och andra leddjur
Mångfotingar, spindlar och insekter har, oberoende av varandra. utvecklat en helt annorlunda typ av exkretionsorgan, malpighiska kärl. De är är blint börjande utbuktningar från den bakre delen av tarmen. De bildar inte primärurin genom filtrering, utan genom sekretion. Den blivande urinen kan modifieras och koncentreras i tarmen, särskilt ändtarmen, innan den lämnar kroppen. Dessa egenskaper gör de malpighiska kärlen till en vattenbesparande evolutionär anpassning till landliv. Eftersom urinen hamnar i tarmen, avger insekter en blandning av avföring och urin.
Hos insekter bildas primärurinen genom aktiv energikrävande sekretion av ämnen, främst kaliumjoner (K+), in i de malpighiska kärlen. Eftersom kärlens hålrum då blir positivt laddade, transporteras negativt laddade kloridjoner (Cl-) sedan passivt in i hålrummet attraherade av den positiva laddningen där. Då stiger osmolariteten ("totalkoncentrationen av lösta ämnen") i hålrummen och blir högre än utanför kärlen. Detta gör att vatten passivt följer efter in i kärlen med osmos. Vattenmängden ökar då i kärlens hålrum. Förenklat uttryckt innehåller hålrummen då en lösning av nästan bara kaliumjoner och kloridjoner, men nästan inga andra lösta ämnen. Många ämnen utanför de malpighiska kärlen kommer då att ha en högre koncentration där än inuti kärlens hålrum. De transporteras då med diffusion in i hålrummen, drivna av sin koncentrationsskillnad.
Många insekter som lever i extremt torr miljö, till exempel mjölbaggar, kan göra urinen extremt koncentrerad i ändtarmen, till den grad att blandningen av avföring och urin avges i närmast fast form. I samband med detta faller urinsyra ut, främst som kristaller av dess kaliumsalt. Detta underlättar urinkoncentreringen. Urinsyra bildas vid nedbrytningen av proteinernas aminosyror och utsöndras i stor mängd.
Insekter som får i sig ett stort överskott av vatten, till exempel bladlöss, kan avge stora mängder mycket utspädd urin. Deras blandning av avföring och urin innehåller till och med en hög halt av det energirika nyttiga ämnet rörsocker (vanligt socker), som de får i sig i överskott när de suger i sig växtsafter. Denna blandning kallas honungsdagg och ätes med förtjusning av många insekter, till exempel myror.
Insekternas malpighiska kärl

De malpighiska kärlen är insekternas utsöndringsorgan. De börjar blint och mynnar mellan mellantarmen ock baktarmen, se bilden ovan. Hos olika arter varierar antalet kärl mellan två och mer än 200. Primärurinen bildas genom sekretion, men mekanismerna är inte fullständigt kända.
Produktionen av primärurin drivs främst av att kaliumjoner (K+) transporteras med aktiv, energikrävande transport in i kärlens inledande avsnitt (se schemat överst till höger på bilden ovan). Energin kommer från molekylen ATP.
När kaliumjoner ansamlas inne i kärlens hålrum uppkommer ett överskott av positiva laddningar där och ett överskott av negativa laddningar utanför. Denna elektriska potentialskillnad driver passivt kloridjoner (Cl-) från utsidan av kärlen in i hålrummet, genom kanaler i cellmembranerna eller på något annat sätt.
När kaliumjoner och kloridjoner (röda och gröna prickar) ansamlas i hålrummet blir osmolariteten ("totalkoncentrationen av lösta ämnen") högre där än utanför. Då drivs vatten passivt in i hålrummet med osmos, oklart i vad mån detta sker direkt genom cellmembranerna, genom vattenkanaler i cellmembranerna eller mellan väggens celler.
Eftersom andra lösta ämnen, förutom kalium och klorid, saknas i hålrummet, blir deras koncentrationer högre utanför de malpighiska kärlen än inuti dem. De transporteras därför passivt in i hålrummet med diffusion, för varje ämne driven av dess koncentrationsskillnad. Alla dessa ämnen betecknas med X och violetta prickar ovan. Olika ämnen tar sig igenom kärlets vägg på olika sätt.
Den genom sekretion bildade primärurinen modifieras sedan med reabsorption och sekretion av många ämnen i de senare delarna av de malpighiska kärlen och i tarmen, särskilt ändtarmen.
Många insekter producerar en koncentrerad hyperosmotisk urin, en evolutionär anpassning till landliv i en mer eller mindre torr miljö. Då absorberas stora mängder vatten i ändtarmen. Detta leder till att urinsyra faller ut främst som kristaller av sitt kaliumsalt. Då sjunker koncentationen av urinsyra och därmed osmolariteten i tarmvätskan, något som underlättar reabsorptionen av vatten med osmos. De flesta insekter bildar urinsyra, inte urinämne, som slutprodukt vid nedbytning av proteinernas aminosyror och måste därför utsöndra stora mängder av detta ämne.
Hos bland annat vissa flugor och hos mjölbaggar finns mycket avancerade mekanismer som återvinner vatten i ändtarmen, så att urinen blir exceptionellt hyperosmotisk. Det handlar om så kallad aktiv motströmsväxling. Den innefattar i princip energikrävande transport av kaliumjoner från ändtarmens hålrum till en kanal, i vilken vätska flödar i motsatt riktning mot flödet i tarmen, och reabsorption av vatten med osmos.
Fördjupning
- Artikeln Hur sker transporter i djurens kroppar? Diffusion och massflöde finns under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten.
- Du hittar artikeln Osmoreglering här nedan.
- Under rubriken Djurens evolution och människans ursprung – Åldrande och livslängd hittar du artikeln Huvudets uppkomst. Om de första bilateralsymmetriska, tvåsidiga, djuren.
- Artikeln Radiär och bilateral symmetri hos djur. Tvåsidiga djur. Varför har vi höger och vänster kroppshalva? finns under rubriken Springa, krypa, kräla, simma, flyga – Hur rör sig olika djur?
- Har myror ett hjärta? Om insekternas öppna cirkulation och människans slutna blodomlopp är en artikel som du hittar under rubriken Hur tickar djurens hjärta? Om blodcirkulation och andning på land och i vatten
- Mer om exkretionsorgan finns här nedanför
- Nästa artikeln nedanför – Grodor i öknen: om utsöndring av ammoniak, urinämne och urinsyra och om vattenbrist i torr miljö
Referenser
- R. C. Brusca och G. J. Brusca: Invertebrates (Sinauer, 2:a upplagan, 2003).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
- K. V. Kardong: The vertebrates (5th ed, McGraw-Hill, 2009).
- E. P. Widmayer, H. Raff, och K. T. Strang: Vander's human physiology (12th ed, McGraw-Hill, 2011).

De flesta groddjur lever som vuxna på land, några av dem dock i sötvatten. De har en fuktig hud, som lätt släpper igenom vatten. De är därför känsliga för uttorkning. De flesta av dem är också beroende av sötvatten för att kunna fortplanta sig. Larverna, grodynglen, lever nästan alltid i sötvatten. Ändå finns det grodor och paddor som anpassat sig till miljöer där vattenbrist är ett stort problem. Det finns faktiskt grodor och paddor som klarar att leva i öknar och andra torra miljöer samt en art som lever i havet. För att förklara hur dessa groddjur klarar ökentorka och saltvatten måste vi först berätta om de slutprodukter efter proteinnedbrytning som djur måste utsöndra från kroppen.
Slutprodukter vid proteinnedbrytning
Nedbrytning av aminosyror från födans och kroppens proteiner leder hos alla djur till att en kvävehaltig slutprodukt bildas. Denna slutprodukt måste djuren göra sig av med. Hos vattendjur är slutprodukten ofta ammoniak, egentligen huvudsakligen ammoniumjoner. Ammoniak är giftigt, men vattendjur kan göra sig av med ammoniaken innan den når farliga koncentrationer. Gälarna utgör nämligen en stor yta som är i direkt kontakt med det omgivande vattnet. Ammoniak eller ammoniumjoner avlägsnas lätt via gälarna. Groddjurens larver, som lever i sötvatten och andas med gälar, har ammoniak som slutprodukt. En del grodor, till exempel klogrodor (Xenopus), lever nästan hela sitt liv i sötvatten. Hos dem bildar även de vuxna grodorna ammoniak, som lätt kan avges till vattnet via huden.
På land måste slutprodukten avges från njurarna via urinen. Om ammoniak hade avgetts via urinen, måste den ha en mycket låg koncentration för att undvika förgiftning. Vattenförlusterna via urinen skulle då bli enormt stora. Eftersom vattenbrist är ett mer eller mindre stort problem för de flesta landdjur, är det därför i stort sett omöjligt för dem att ha ammoniak som slutprodukt. Fåglar och de flesta andra kräldjur avger urinsyra, däggdjur avger urea (urinämne), se bilden nedan.

Notera att ammoniak innehåller en kväveatom, urea två och urinsyra fyra. Lösta molekyler kan hålla kvar eller dra in vatten i urinen genom så kallad osmos. Denna osmotiska effekt, mätt per kväveatom, är störst för ammoniak, mindre för urea och minst för urinsyra, något bidrar till att göra urea och urinsyra vattenbesparande
Hur groddjur klarar landliv: urea och vattenupptag
För att spara på vatten har landlevande grodor och paddor nästan alltid urea som slutprodukt efter aminosyranedbrytningen, precis som däggdjuren. Urea är inte särskilt giftigt och kan därför avges i koncentrerad form med urinen. Vattenförlusterna via urinen blir då små. Grodor och paddor kan dessutom ofta tolerera höga ureakoncentrationer i vävnaderna. De kan även använda utspädd urin som lagras i urinblåsan som ett vattenförråd. Till skillnad från däggdjur kan de nämligen absorbera vatten från urinblåsan. På land kan de också ta upp vatten genom huden. De pressar då kroppen mot ett fuktigt underlag, till exempel jord eller mossa, och absorberar vatten från underlaget genom huden via osmos. Osmos innebär att vatten förflyttar sig från en lösning med låg total halt av lösta ämnen till en med hög, alltså från en utspädd vattenlösning till en mera koncentrerad. Regnvatten och annat vatten på markytan har en betydligt lägre totalhalt av lösta ämnen än groddjurens kroppsvätskor.

Hur groddjur klarar landliv: att undvika uttorkning
Det finns grodor och paddor som lever i öknen trots att de har en vattengenomsläpplig hud. Dessa djur har beteenden som gör att de undviker uttorkning. De är ofta nattdjur. De gömmer sig gärna i hålor i marken där temperaturen är lägre och luftfuktigheten högre. När det behövs använder de urinblåsan som vattenförråd. Flera av dem överlever torrperioder nergrävda i marken. En del av dem är då omgivna av en skyddande kokong bestående av avkastad hud. I nergrävt tillstånd kan de ackumulera stora mängder urea utan att ta skada. Ureakoncentrationen hos en del av dem kan till och med bli så hög att de tar upp vatten från marken utanför kokongen genom osmos. När regn fallit i öknen, kryper de upp ur sina hålor och fortplantar sig oerhört snabbt. När öknen torkar ut igen, har nästan generation grodor redan blivit vuxna. De kan krypa ner i marken och där invänta nästa regnperiod.
En havslevande groda. Grönfläckig padda
De anpassningar som de ökenlevande groddjuren uppvisar har också möjliggjort ett liv i eller vid havet. Problemet med havsliv för ett groddjur är nämligen till stor del att det salta eller bräckta vattnet är mer koncentrerat än djurets kroppsvätskor. Djuret förlorar alltså vatten genom osmos via huden. Det handlar med andra ord om uttorkning. En hög ureahalt kan innebära att djuret kommer i osmotisk jämvikt med vattnet eller till och med att det tar upp vatten genom sin hud. Den utrotningshotade grönfläckiga paddan (Pseudepidalea viridis) lever i Sverige på havsstränder med brackvattensamlingar och klarar sig fint, bland annat tack vare sin förmåga att ansamla urea. Den berömda krabbätande grodan i Sydostasien (Fejervarya cancrivora) lever i mangroveträsk. Det är det enda groddjur som kan leva i normalt, obräckt havsvatten. Även den förlitar sig på sin förmåga att ansamla urea i kroppsvätskorna. Den kan till och med ta upp vatten via huden genom osmos. Den behöver därför inte dricka av havsvattnet.
Grodor med vattentätad hud
En del grodor och paddor har ett extra tjockt hornlager i huden. Detta gör att huden släpper igenom mindre vatten, men den är ändå avsevärt mera vattengenomsläpplig än huden hos kräldjur, inklusive fåglar, och hos däggdjur. Men det finns några trädlevande tropiska grodor som har gått ett par steg längre i sin anpassning till vattenbrist. Deras hud är ungefär lika ogenomsläpplig för vatten som huden hos många kräldjur. Orsaken är att huden är täckt av ett lager av fettämnen, huvudsakligen vaxer, som är starkt vattenskyende. Arter av det amerikanska släktet Phyllomedusa (se bilden överst på sidan och videorna nedan) gnider in sig med vax från hudkörtlar med hjälp av frambenen. Det ser ut ungefär som när människor gnider in sig med solskyddsmedel på stranden.
Värt att notera är att huden hos kräldjur, inklusive fåglar, och hos däggdjur inte vattentätas av fett på hudens yta. Däggdjurens talgkörtlar mynnar i hårsäckarna. Talgens funktion är framför allt att göra pälsens hårstrån vattenskyende och i någon mån att mjuka upp huden. Huden vattentätas i stället av skikt av fettämnen mellan de döda cellerna inuti hudens översta lager, hornlagret. Forskare har länge trott att fåglar vattentätar sin fjäderdräkt genom att gnida in den med sekret från den så kallade gumpkörteln. Nyare studier har ifrågasatt detta. Kanske är det framför allt fjädrarnas struktur som gör dem vattenskyende.
Grodor som avger urinsyra i stället för urea
De tropiska grodorna inom släktena Phyllomedusa och Chiromantis har en annan intressant vattenbesparande anpassning. De har inte, som de flesta andra vuxna groddjur, urea som slutprodukt vid aminosyranedbrytningen. De har i stället urinsyra, precis som fåglar, de flesta övriga kräldjur och de flesta insekter. Fördelen med urinsyra är att den är dåligt vattenlöslig och därför kan fällas ut som kristaller av urater, urinsyrans salter. När detta sker i den blivande urinen, späs den ut, mer vatten kan återvinnas från den genom osmos och en mindre mängd vatten förloras via den slutliga urinen.
Referenser
- N. Castillo: Breviceps macrops (AmphibiaWeb, uppdaterad 2011-12-20).
- R.W. Hill, G.A. Wyse, och M. Anderson: Animal physiology (3rd ed, Sinauer, 2012).
- V. Mercurio, W. Böhme, B. Streit: Reproductive diversity of Malawian anurans (Herpetology Notes 2:175-183, 2009).
- K. Schmidt-Nielsen: Animal physiology (5th ed, Cambridge University Press, 1997).
- P. Willmer, G. Stone, och I. Johnston: Environmental physiology of animals (2nd ed, Blackwell, 2005).

Det finns många sätt att släcka törsten utan att dricka. En del djur kan ta upp flytande vatten genom huden. Andra kan ta upp vattenånga i gasform från luften. Det finns till och med djur som till stor del klarar sig på vatten som de själva producerar.
Många grodor och andra amfibier dricker inte alls, utan absorberar i stället vatten genom huden. De behöver inte ens vara nedsänkta i vatten, utan det räcker med att de trycker sig mot fuktig jord eller något annat fuktigt underlag. Hastigheten som de kan absorbera vatten med beror på hudens genomsläpplighet för vatten och på skillnaden i vattenpotential mellan grodans kroppsvätskor och underlaget. Vattenpotentialen är ett mått på ett materials benägenhet att dra till sig vatten. Uttorkad groda och vattenrik jord gör att vattnet rör sig in i grodan. Vattnet transporters in i grodan genom så kallad osmos, beroende på att totalhalten av lösta ämnen är högre i grodans kropp än i underlaget. Grodan kan också öka sin huds genomsläpplighet för vatten när det behövs.
Vid utflykter bort från sin fuktiga hemvist tar en del grodor med sig reservvatten i form av utspädd urin i urinblåsan. De återanvänder vattnet i urinen genom att öka genomsläppligheten för vatten i urinblåsans vägg. Eftersom totalhalten av lösta ämnen är större i grodans kroppsvätskor än i den utspädda urinen, absorberar grodan vatten från urinen med osmos. Salt, i form av natriumjoner och kloridjoner, transporteras också ut från blåsan med en aktiv energikrävande process. Då följer vatten efter med osmos. Grodorna kan fylla sin urinblåsa med mycket vatten, om de ska vara borta länge i varmt och torrt väder.

En del insekter och andra leddjur, till exempel dammkvalstret, kan absorbera vattenånga som finns i luften. Det handlar om gasformigt vatten i en luft, som inte är mättad med vattenånga. Det rör sig alltså inte om kondensering av vattendroppar ur luft som är övermättad på vattenånga. Även i torra miljöer finns det en del vattenånga i luften. Forskare har inte lyckats klarlägga, hur dessa leddjur kan ta upp vattenångan. En del av dem producerar kanske en mycket koncentrerad saltlösning i munnen eller ändtarmen. En sådan saltlösning kan ta upp vattenånga från luft, som inte är vattenmättad. De leddjur som absorberar vattenånga får inte i sig så stora mängder vatten. Men de förlorar I gengäld inte så mycket vatten. Deras yttre skelett, kutikulan, är nämligen täckt av ett vaxlager, som har en mycket låg genomsläpplighet för vatten. Dessutom kan dessa djur producera en mycket koncentrerad urin. Därmed sparar de på vatten, som annars hade behövts till att föra bort avfallsprodukter.
Vissa ökenlevande ödlor och fåglar behöver inte heller dricka för att få i sig vatten utan det räcker med att äta växter som innehåller mycket vatten, till exempel suckulenta växter med köttiga stammar och blad. Dessa djur sparar också mycket vatten genom att avge urinen i nästan fast form.
Det finns små ökengnagare som kan överleva på ;metabolt vatten tillsammans med de små vattenmängder som finns i den torra växtföda de äter. Metabolt vatten är det vatten som kroppen själv producerar vid förbränningen av kolhydrater, fett och proteiner från födan eller från kroppens energiförråd. En människa producerar några deciliter metabolt vatten per dygn. Ökengnagarna måste vara mycket sparsamma med kroppens vatten för att kunna klara sig på en så liten vattentillförsel. Det kan de vara bland annat genom att avge en mycket koncentrerad urin och en torr avföring. De är ofta aktiva på natten, då ökenluften är svalare och fuktigare än på dagen. Dagarna tillbringar de i sina svala och fuktiga jordhålor. De minskar vattenförlusterna via utandningsluften genom att återvinna vatten i nosen.
Referenser
R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).
K. Schmidt-Nielsen: Animal physiology (5:e upplagan, Cambridge University Press, 1997).

Vi människor är beroende av tillgång till sötvatten att dricka för att kunna överleva. Detta faktum är dock inte allmängiltigt i hela djurvärlden. Många arter kan tillfredsställa sina vattenbehov genom att dricka saltvatten direkt från havet. Om vi människor hade gjort detta, hade vi förvärrat vår situation. Många skeppsbrutna sjömän har blivit så törstiga att de druckit havsvatten, vilket lett till att de avlidit av saltförgiftning. Hur kan detta komma sig? Svaret ligger i olika förmåga att koncentrera saltet då det skall avges från kroppen.
För att kunna tillgodogöra sig saltvatten krävs att du kan göra dig av med saltet i en mindre volym vatten än det intagits i, det vill säga salterna måste kunna koncentreras. Vår främsta väg att göra oss av med salt är via urinen. Genom njurarna kan vi producera en urin som är mer koncentrerad än våra övriga kroppsvätskor, men endast till en viss gräns. Vi kan ej producera en urin som är saltare än havsvatten.
De joner som finns i högst koncentration i havsvatten är natriumjoner och kloridjoner, samma joner som ingår i koksalt. Även magnesiumjoner och sulfatjoner finns i ganska höga koncentrationer i havsvatten. Maximalt koncentrerad urin från en människa innehåller lägre koncentrationer av natriumjoner och kloridjoner än havsvatten. Dricker vi en liter havsvatten och avger en liter urin har vi således inte tillfört kroppen något vatten. I stället har vi tillfört salt och blivit saltförgiftade. Till detta kommer att havsvatten har en laxerande effekt, vilket leder till extra vattenförluster i form av diarréer. Magnesiumjoner och sulfatjoner ingår i så kallat epsomsalt som har en laxerande effekt.
Fåglar och reptiler som lever i havsmiljöer har sämre förmåga än vi människor att koncentrera sin urin, men kan ändå dricka det salta havsvattnet. Evolutionen har försett dessa arter med så kallade saltkörtlar som är placerade i huvudregionen. Hos fåglar är de placerade ovanför ögongloberna och töms via näshålan. Körtlarna producerar en mycket koncentrerad saltlösning, mer koncentrerad än havsvattnet avseende natriumjoner och kloridjoner. Tack vare dessa körtlar behöver inga stora mängder urin avges och större delen av det vatten som intagits kan behållas av kroppen och användas till livsnödvändiga processer.
Även benfiskar som lever i saltvatten har utvecklat metoder för att kunna dricka vattnet de lever i. Till skillnad från sötvattenslevande fiskar, måste dessa fiskar dricka vatten. Miljön de lever i är mer koncentrerad avseende salter och andra ämnen än deras kroppsvätskor. Detta innebär att de förlorar vatten till omgivningen genom osmos. Deras urin är inte mer koncentrerad än kroppsvätskorna, men de kan i stället avge saltet genom gälarna och hushållar på så sätt med vattnet.
Om vi människor befinner oss i en situation där vi upplever stor törst, men endast har tillgång till mycket salt vatten är det följaktligen betydligt bättre att inte dricka alls. Om vattnet däremot har ungefär lika hög koncentration som den våra njurar kan producera i urinen, skulle det kanske vara en fördel att dricka vattnet. Kroppen får inget mer vatten, men den producerade urinen kan föra med sig urinämne och andra slutprodukter från kroppen.
Referenser
R.W. Hill, G.A. Wyse, och M. Anderson: Animal Physiology (3rd ed, Sinauer, 2012).

Fråga en biolog
Hittar du inte svaret på dina frågor? Titta in på vår blogg och ställ din fråga.
Författare
Anders Lundquist
Anders Lundquist har varit verksam på institutionen i mer än 35 år. Han har undervisat i framförallt djurfysiologi och människans fysiologi. Han har handlett många studenter genom åren och fått studentkårens pedagogiska pris. I mer än tjugo år har han drivit en populärvetenskaplig webbplats om djur med artiklar och frågelåda. Det är därifrån de här artiklarna kommer.
Vissa artiklar är skrivna av studenter, men de har faktagranskats, redigerats och uppdaterats av Anders Lundquist.
Bilder
Bilderna på den här sidan kommer från ett antal kreatörer som har släppt sina alster under Creative Commons licenser. Vill du veta mer om vilka kreatörerna, webbplatserna och licenserna är kan du titta på sidan där vi listar dem.